ブテルア・グラシリス
| ブルーグラマ | |
|---|---|
 | |
| 科学的分類 | |
| 王国: | 植物界 |
| クレード: | 維管束植物 |
| クレード: | 被子植物 |
| クレード: | 単子葉植物 |
| クレード: | ツユクサ類 |
| 注文: | ポアレス |
| 家族: | イネ科 |
| 亜科: | クロリドイデア科 |
| 属: | ブテロア |
| 種: | B. gracilis |
| 二名法名 | |
| ブテルア・グラシリス | |

{kind=link}
ブテロウア・グラシリス(ブルーグラマ)は、北アメリカ原産の長寿命の暖季型( C4 )多年草で ある。 [ 2 ] [ 4 ] [ 5 ]
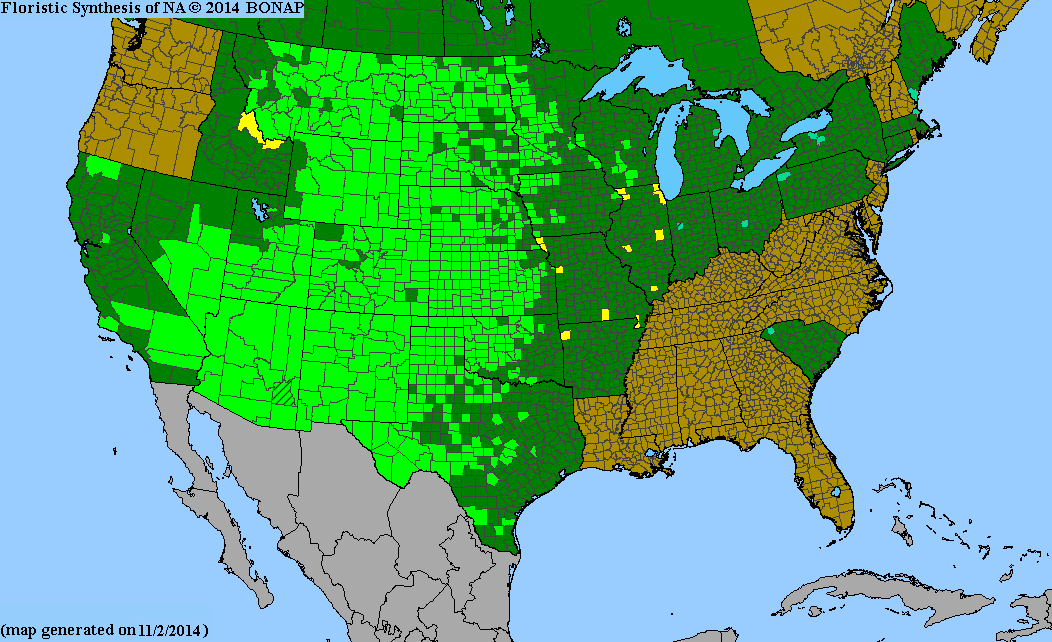
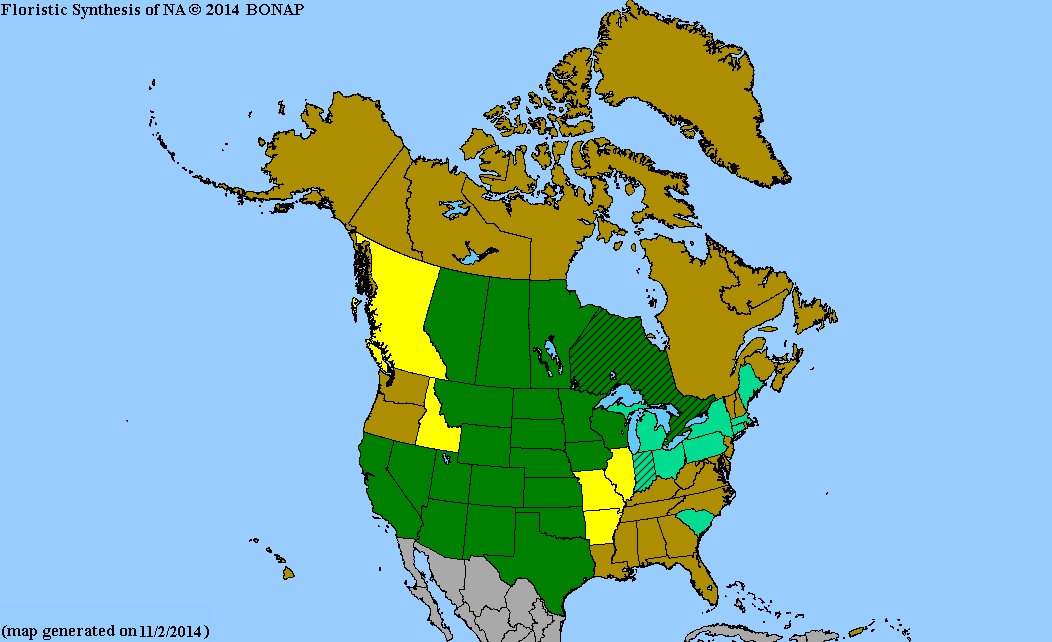
最も一般的に見られるのは、カナダのアルバータ州から東はマニトバ州、南はロッキー山脈、グレートプレーンズ、米国中西部諸州を越えて、メキシコのメキシコ高原北部にかけてです。
ブルーグラマは、グレートプレーンズ中央部および南部の短草プレーリーにおける純一次生産の大部分を占めています。これは緑色または灰色がかった草で、背丈が低く、乾燥に強く、手入れの手間もほとんどかかりません。[ 6 ]
説明
{kind=link}

{kind=link}

{kind=link}

_II.jpg){kind=link}
_II.jpg/1280px-Bouteloua_gracilis_(Poaceae)_II.jpg)
ブルーグラマは、緑色から灰色がかった葉を持ち、幅は3mm(0.1インチ)未満、長さは25~250mm(1~10インチ)です。成熟すると、全体の高さは15~30cm(6~12インチ)になります。[ 7 ]
花茎(稈)は長さ7~18インチ(18~46cm)です。先端には1~4個(通常は2個)の櫛状の花穂があり、 [ 8 ]花穂は花茎から鋭角に伸びています。[ 7 ]各花穂には20~90個の小穂があります。[ 7 ]各小穂は長さ5~6mm(0.20~0.24インチ)で、稔性の小花が1つと、縮小した不稔性の小花が1~2つあります。[ 9 ]小穂の下には2つの穎花があり、1つは長さ1.5~3mm(0.06~0.12インチ)で、もう1つは長さ3.5~6mm(0.14~0.24インチ)です。[ 9 ]稔性の小花は長さ5~5.5mm(0.20~0.22インチ)の外套(苞葉)があり、先端に3本の短い芒(剛毛)があり、不稔性の小花は長さ約2mm(0.08インチ)の外套と長さ約5mm(0.2インチ)の3本の芒があります。[ 8 ] [ 10 ]受粉すると、稔性の小花は長さ2.5~3mm(0.10~0.12インチ)の長楕円形の褐色の種子を生成します。[ 11 ]種子が成熟すると、小穂全体が分離しますが、2つの穎果は残ります。
根は通常、外側に12~18インチ(30~46cm)、深さ3~6.5フィート(0.9~2.0m)成長します。[ 7 ]
ブルーグラマは種子から容易に定着するが、分げつによる栄養繁殖に大きく依存する。種子生産は遅く、土壌水分と温度に依存する。風によって散布された種子は数メートル(6フィート)しか到達せず、昆虫、鳥、哺乳類が散布媒介となってそれ以上の距離まで到達する。実生の定着、生存、成長は、実生の根域の水分を効率的に利用する近隣の成体植物から隔離された状態で最大限に促進される。定着を成功させるには、不定根の伸長と発達の間に適度な土壌水分が必要である。 [ 7 ]
定着した植物は放牧、寒さ、干ばつに耐性があるが、長期の干ばつは根の数と広がりの減少につながる。彼らは機会主義的な水利用戦略を採用しており、利用可能な水があれば迅速に利用し、条件が劣る場合は休眠状態に入る。遷移の観点から見ると、ブルーグラマは極相種への遷移が遅い。撹乱後の回復は遅く、撹乱の種類と程度に依存する。[ 7 ]
分布
ブルーグラマは、グラマ科の草本植物の中で最も広く分布しています。ほとんどの土壌で生育し、地域の条件に容易に適応します。北はアルバータ州、南はメキシコまで分布しています。アメリカ合衆国中西部の大部分に分布し、東はミズーリ州とテキサス州、西は南カリフォルニアまで広がっています。東部の一部の州や南アメリカにも導入されています。 [ 12 ]
園芸と農業
ブルーグラマは飼料として重宝されており、最も貴重なグラマです。南西部では理想的な牧草地です。過度の放牧や極度の干ばつにも耐え、家畜にとっても非常に食べやすいです。[ 12 ]ブルーグラマの放牧地では、望ましくない雑草の侵入を防ぐ効果もあるかもしれません。[ 13 ]
ブルーグラマは園芸産業で栽培されており、多年草庭園、自然派および在来植物 を用いた造園、生息地の復元プロジェクト、住宅 地、公共施設、高速道路の浸食防止などに利用されています。ブルーグラマの花はドライフラワーアレンジメントにも用いられます。ブルーグラマはコロラド州とニューメキシコ州の州草です。イリノイ州では絶滅危惧種に指定されています。[ 2 ]
ブルーグラマは他のグラマ科植物に比べてゲノムが大きい。[ 14 ]二倍体または四倍体である。[ 15 ]
ズニ族は、草の束を束ね、切り取った方の端をヘアブラシとして、もう一方の端をほうきとして使います。また、束はヤギの乳を濾すのにも使われます。[ 16 ]コスタノ族、あるいはオローニ族は、中空の茎をストローとして使っていました。[ 17 ]ナバホ族は、それを羊や馬の飼料として使っていました。[ 18 ]
生態学
ブルーグラマは、米国の短草ステップ生態地域の優占種である。[ 19 ] [ 20 ]大平原に生息するブルーグラマの個体群は遺伝的に分化しており(小さな空間スケールでも)、気候に関連した機能的形質の変異を示している。 [ 21 ] [ 22 ] [ 15 ]より乾燥した草原に生息するブルーグラマは、表現型の可塑性が高いことも特徴である。[ 21 ]
これは、ガリタスキッパーリング、ミドリスキッパー、パハスカスキッパー、アカゲザルスキッパー、シミウスロードサイドスキッパー、ウンカススキッパーの幼生宿主である。[ 23 ]
参考文献
- ^ NatureServe (2023). 「Bouteloua gracilis Blue Grama」 . NatureServe Explorer . バージニア州アーリントン: NatureServe . 2023年4月28日閲覧。
- ^ a b c NRCS . 「Bouteloua gracilis」 . PLANTSデータベース.米国農務省(USDA).
- ^ 「Bouteloua gracilis」 .国際植物名索引(IPNI) .キュー王立植物園、ハーバード大学植物標本館・図書館、オーストラリア国立植物園.
- ^ Kartesz, John T. (2014). 「Bouteloua gracilis」 .北米植物アトラス(NAPA)の郡レベル分布図. 北米生物相プログラム(BONAP) . 2018年7月13日閲覧。
- ^ Kartesz, John T. (2014). 「Bouteloua gracilis」 .北米植物アトラス(NAPA)による州レベル分布図. 北米生物相プログラム(BONAP) . 2018年7月13日閲覧。
- ^ Smoliak, S.; Ditterline, RL; Scheetz, JD; Holzworth, LK; Sims, JR; Wiesner, LE; Baldridge, DE; Tibke, GL 「ブルーグラマ(Bouteloua gracilis)」。モンタナ州政府機関植物資料ハンドブック。モンタナ州立大学エクステンションサービス。2010年6月10日時点のオリジナルよりアーカイブ。
- ^ a b c d e fアンダーソン、ミシェル・D. (2003). 「Bouteloua gracilis」 .火災影響情報システム (FEIS) . 米国農務省 (USDA), 森林局 (USFS), ロッキー山脈研究ステーション, 火災科学研究所. 2016年8月24日閲覧。
- ^ a b Sun、Bi-xing;フィリップス、シルビア M. 「ブーテロア・グラシリス」。中国の植物相。 Vol. 22 – eFloras.org、ミズーリ州セントルイスのミズーリ植物園およびマサチューセッツ州ケンブリッジのハーバード大学ハーバリア経由。
- ^ a bヒルティ、ジョン (2020). 「ブルーグラマ(Bouteloua gracilis)」イリノイ野生の花.
- ^孫碧興;フィリップス、シルヴィア M. 「ブーテロア」。中国の植物相。 Vol. 22 – eFloras.org、ミズーリ州セントルイスのミズーリ植物園およびマサチューセッツ州ケンブリッジのハーバード大学ハーバリア経由。
- ^チャイカ、ケイティ;ピーター・ジウク (2016)。「ブーテロア・グラシリス(ブルーグラマ)」。ミネソタ州の野生の花。
- ^ a bグールド、フランク・W. (1951). 『アメリカ南西部のイネ科植物』アリゾナ大学ツーソン校. p. 146.
{{cite book}}:|work=無視されました (ヘルプ) - ^ポレンスキー, ローレン・M.; ダーナー, ジャスティン・D.; オーガスティン, デイビッド・J.; ミルチュナス, ダニエル・G. (2017). 「短草ステップにおける75年間の持続的放牧強度処理後の植物群集構成」. Rangeland Ecology & Management . 70 (4): 456– 464. doi : 10.1016/j.rama.2016.12.001 . hdl : 10150/667452 . S2CID 90458063 .
- ^ベネット、M (2000). 「被子植物の核DNA量とその現代的利用 ― 807の新たな推定値」 Annals of Botany . 86 (4): 859– 909. doi : 10.1006/anbo.2000.1253 .
- ^ a b Butterfield, Bradley J.; Wood, Troy E. (2015). 「Bouteloua gracilis (Poaceae) における機能的形質の変異は、倍数性ではなく、地域気候と栽培によって予測される」. Plant Ecology . 216 (10): 1341– 1349. doi : 10.1007/s11258-015-0510-8 . ISSN 1385-0237 . S2CID 18031688 .
- ^スティーブンソン、マチルダ・コックス(1915年)「ズニ族インディアンの民族植物学」SI-BAE年次報告書第30号、p.83。
- ^ Bocek, Barbara R. (1984). 「ジョン・P・ハリントンのコレクションに基づく、カリフォルニアのコスタノアン・インディアンの民族植物学」 . Economic Botany . 38 (2): 255. doi : 10.1007/BF02858839 . S2CID 21852804 .
- ^エルモア、フランシス・H. (1944). 『ナバホ族の民族植物学』サンタフェ、ニューメキシコ州:アメリカン・リサーチ・スクール、p. 25.
- ^ウィリアム・K・ラウエンロス、イングリッド・C・バーク編 (2008).短草ステップの生態学:長期的な視点. オックスフォード大学出版局. ISBN 978-0-19-972280-8. OCLC 690360063 .
{{cite book}}:|last=一般的な名前があります(ヘルプ)CS1 maint: 複数の名前: 著者リスト (リンク) - ^ Munson, Seth M.; Lauenroth, William K. (2009). 「短草ステップにおける優占種の除去に対する植物個体群と群集の反応:優占種の除去に対する植物の反応」. Journal of Vegetation Science . 20 (2): 224– 232. doi : 10.1111/j.1654-1103.2009.05556.x .
- ^ a b Hoffman, Ava M.; Bushey, Julie A.; Ocheltree, Troy W.; Smith, Melinda D. (2020). 「地域および地方スケールにおける遺伝的・機能的変異は、草原の基盤となる草地における気候と関連している」 . New Phytologist . 227 (2): 352– 364. doi : 10.1111/nph.16547 . ISSN 0028-646X . PMID 32176814 .
- ^ Smith, Steven E.; Arredondo, Tulio; Aguiar, Martín; Huber-Sannwald, Elisabeth ; Alpuche, Angel; Aguado, Armando; Grageda, Oscar A.; Halbrook, Kandres; Bottini, Cecilia (2009). 「3つの環境における多年生イネ科植物の微細空間遺伝構造」. Rangeland Ecology & Management . 62 (4): 356– 363. doi : 10.2111/08-159.1 . hdl : 10150/643040 . S2CID 44199283 .
- ^ The Xerces Society (2016)、「蝶のためのガーデニング:美しく有益な昆虫を引き寄せ、守る方法」、Timber Press。
{kind=link}
{kind=link}
外部リンク
