| オルトミクソウイルス科 | |
|---|---|

| |
| インフルエンザA型およびインフルエンザB型ウイルスのゲノム、mRNA、およびビリオンの図 | |
| ウイルスの分類 | |
| (ランク外): | ウイルス |
| レルム: | リボビリア |
| 王国: | オルタナウイルス科 |
| 門: | ネガルナビリコタ |
| クラス: | インストビリセテス |
| 注文: | アーティキュラウイルス科 |
| 家族: | オルトミクソウイルス科 |
| 属 | |
|
本文参照 | |
オルトミクソウイルス科(古代ギリシャ語 のὀρθός (orthós) 「まっすぐな」と μύξα (mýxa) 「粘液」に由来) [1]は、マイナスセンス RNAウイルス科です。アルファインフルエンザウイルス、ベタインインフルエンザウイルス、、デルタインフルエンザウイルス、イサウイルス、 、クアランジャウイルス、サル ディノウイルス、トゴトウイルスの9つの属が含まれます。最初の4つの属には、鳥類(鳥インフルエンザも参照)およびヒトを含む哺乳類にインフルエンザを引き起こすウイルスが含まれます。イサウイルスはサケに感染し、トゴトウイルスはアルボウイルスで、脊椎動物と無脊椎動物(ダニや蚊など)に感染します。 [2] [3] [4]クアランジャウイルスもアルボウイルスであり、脊椎動物(鳥類)と無脊椎動物(節足動物)に感染します。
脊椎動物に感染するインフルエンザウイルスには、核タンパク質とマトリックスタンパク質の抗原性の違いによって識別される 4 つの属があります。
- アルファインフルエンザウイルスは、ヒト、他の哺乳類、鳥類に感染し、あらゆるインフルエンザのパンデミックを引き起こす。
- ベタインインフルエンザウイルスはヒトとアザラシに感染する
- ガンマインフルエンザウイルスはヒトと豚に感染する
- デルタインフルエンザウイルスは豚や牛に感染します。
構造

インフルエンザウイルスのビリオンは多形性であり、ウイルスエンベロープは球状と糸状の形態をとる。一般的に、ウイルスの形態は楕円形で粒子径は100~120 nm、糸状で粒子径は80~100 nm、長さは最大20 μmである。[5]エンベロープには約500個のスパイク状表面突起があり、それぞれ表面から10~14 nm突出し、表面密度は様々である。主要な糖タンパク質(HA)スパイクは、ノイラミニダーゼ(NA)スパイクのクラスターによって不規則に挟まれており、HAとNAの比は約10対1である。[6]
ウイルスエンベロープは脂質二重膜で構成され、糖タンパク質のスパイクが固定されており、ヌクレオカプシド(様々なサイズの核タンパク質)を包んでいる。ウイルス粒子内の配置は不明である。リボ核タンパク質は糸状で、長さは50~150nm、らせん対称性を有する。[7]
ゲノム

オルソミクソウイルス科のウイルスは、6~8個の分節からなる直鎖状のマイナスセンス一本鎖RNAを含み、ゲノム全長は10,000~14,600ヌクレオチド(nt)です。[7]例えば、インフルエンザAのゲノムは、8個の分節化されたマイナスセンスRNA(合計13.5キロベース)から構成されています。[8]
インフルエンザウイルスタンパク質の中で最も特徴的なのは、ウイルス粒子の外側に存在する2つの大きな糖タンパク質、ヘマグルチニンとノイラミニダーゼです。ヘマグルチニンは、ウイルスが標的細胞に結合し、ウイルスゲノムが標的細胞に侵入するのを媒介するレクチンです。 [9]一方、ノイラミニダーゼは、成熟したウイルス粒子に結合する糖を切断することで、感染細胞から子孫ウイルスを放出する酵素です。ヘマグルチニン(H)とノイラミニダーゼ(N)タンパク質は、抗体や抗ウイルス薬の重要な標的であり、[10] [11] A型インフルエンザウイルスの異なる血清型を分類するために使用され、 H5N1のHとNはここから来ています。
ゲノム配列には末端反復配列があり、これらは両端(すなわち5'末端と3'末端の両方)で繰り返されている。5'末端の末端反復配列は12~13ヌクレオチド長である。3'末端のヌクレオチド配列は同一であり、同じファミリーの属間では同一であり、RNA(セグメント)の大部分、またはすべてのRNA種で同一である。3'末端の末端反復配列は9~11ヌクレオチド長である。カプシド化された核酸は完全にゲノム核酸である。各ウイルス粒子には欠陥のある干渉コピーが含まれる可能性がある。インフルエンザA(特にH1N1)では、PB1の選択的読み枠からPB1-F2が生成される。M遺伝子とNS遺伝子は、選択的スプライシングによってそれぞれ2つの遺伝子(合計4つの遺伝子)を生成する。[12]
複製サイクル

典型的には、インフルエンザは感染した哺乳類の咳やくしゃみによって空気感染し、ウイルスを含むエアロゾルを発生させるほか、感染した鳥類の糞便を介して感染する。また、唾液、鼻汁、糞便、血液によっても感染する。感染はこれらの体液や汚染された表面との接触によって起こる。特定の表面(すなわち宿主の体外)においては、インフルエンザウイルスは人体温で約1週間、0℃(32℉)では30日以上、極低温(北東シベリアの湖沼など)では無期限に感染力を維持する。インフルエンザウイルスは消毒剤や洗剤によって容易に不活化される。[13] [14] [15]
ウイルスは、その表面のヘマグルチニン糖タンパク質と相互作用して、宿主の表面、特に肺や喉の上皮細胞の表面にあるシアリン酸糖に結合します(感染図のステージ1)。 [16]細胞はエンドサイトーシスによってウイルスを輸入します。エンドソームの酸性pH環境では、ヘマグルチニンタンパク質の一部がウイルスエンベロープを液胞の膜に融合し、ウイルスRNA(vRNA)分子、アクセサリタンパク質、RNA依存性RNAポリメラーゼを宿主細胞の細胞質に放出します(ステージ2)。[17]これらのタンパク質とvRNAは複合体を形成し、宿主細胞の核に輸送され、そこで宿主自身のRNA依存性RNAポリメラーゼが相補的なプラスセンスcRNAの転写を開始します(ステップ3aとb)。[18] cRNAは細胞質へ輸送されて翻訳されるか(ステップ4)、宿主核内に留まる。新たに合成されたウイルスタンパク質は、ゴルジ体を介して宿主細胞表面に分泌されるか(ノイラミニダーゼとヘマグルチニンの場合、ステップ5b)、宿主核へ輸送され、そこでvRNAと結合して新たなウイルスゲノム粒子を形成する(ステップ5a)。他のウイルスタンパク質は、宿主細胞内で複数の作用を有し、例えば細胞内mRNAを分解し、その結果放出されたヌクレオチドをvRNA合成に利用したり、宿主細胞のmRNAの翻訳を阻害したりする。[19]
ウイルス粒子は、マイナス鎖vRNA(新しく生成されたウイルスのゲノムを形成)、RNA依存性RNA転写酵素、およびその他のウイルスタンパク質から構成される。ヘマグルチニンおよびノイラミニダーゼ分子は、宿主細胞膜の隆起部にクラスターを形成する。vRNAおよびウイルスコアタンパク質は核から出て、この膜突起部に入る(ステップ6)。成熟したウイルスは、宿主のリン脂質膜の球体内で宿主細胞から出芽し、この膜コートとともにヘマグルチニンおよびノイラミニダーゼを獲得する(ステップ7)。[20]前と同様に、ウイルスはヘマグルチニンを介して同じ宿主細胞のカプセルに付着する。成熟ウイルスは、ノイラミニダーゼが宿主細胞からシアリン酸残基を切断すると、分離する。[16]新しいインフルエンザウイルスが放出されると、宿主細胞は死に、他の宿主細胞で感染が繰り返される。

オルソミクソウイルス科ウイルスは、核内で複製する2種類のRNAウイルスのうちの1つです(もう1つはレトロウイルス科)。これは、オルソミクソウイルスの機構が独自のmRNAを生成できないためです。これらのウイルスは、キャップスナッチングと呼ばれるプロセスで、細胞内のRNAをプライマーとして使い、ウイルスのmRNA合成を開始します。[21]核内に入ると、RNAポリメラーゼタンパク質PB2が細胞内のプレmRNAを見つけて、その5′キャップ末端に結合します。次に、RNAポリメラーゼPAが細胞内のmRNAを5′末端付近で切断し、このキャップ断片をプライマーとして使い、残りのウイルスRNAゲノムをウイルスmRNAに転写します。[22]これは、mRNAが細胞のリボソームに認識されて翻訳されるためには、5′キャップが必要なためです。
RNA校正酵素が存在しないため、RNA依存性RNA転写酵素は、インフルエンザvRNAのおおよその長さである約1万ヌクレオチドごとに1つのヌクレオチド挿入エラーを起こします。そのため、ほぼすべての新しく製造されたインフルエンザウイルスは、ゲノムに変異を含みます。[23]ゲノムが8つのvRNAセグメントに分かれているため、複数のインフルエンザウイルスが同じ細胞に感染した場合(重複感染)、遺伝子の混合(再集合)が可能になります。ウイルスの子孫にパッケージ化されたゲノムセグメントの変化は、新しい行動、時には新しい宿主種に感染する能力や、宿主集団の古いゲノムに対する防御免疫を克服する能力(この場合は抗原シフトと呼ばれます)を与えます。[10]
分類
系統分類学に基づくRNAウイルスの分類では、マイナス鎖RNAウイルスというサブカテゴリが含まれ、これにはアルティキュラウイルス目とオルトミクソウイルス科が含まれる。この科には以下の属が含まれる。[24]
- アルファインフルエンザウイルス
- ベタインインフルエンザウイルス
- デルタインフルエンザウイルス
- ガンマインフルエンザウイルス
- イサウイルス
- マイキスウイルス
- クアランジャウイルス
- サルディノウイルス
- トゴトウイルス
インフルエンザの種類
インフルエンザウイルスには4つの属があり、それぞれに1つの種(型)のみが含まれます。インフルエンザA型とC型は様々な種(ヒトを含む)に感染しますが、インフルエンザB型はほぼヒトにのみ感染し、インフルエンザD型は牛と豚に感染します。[25] [26] [27]
インフルエンザA
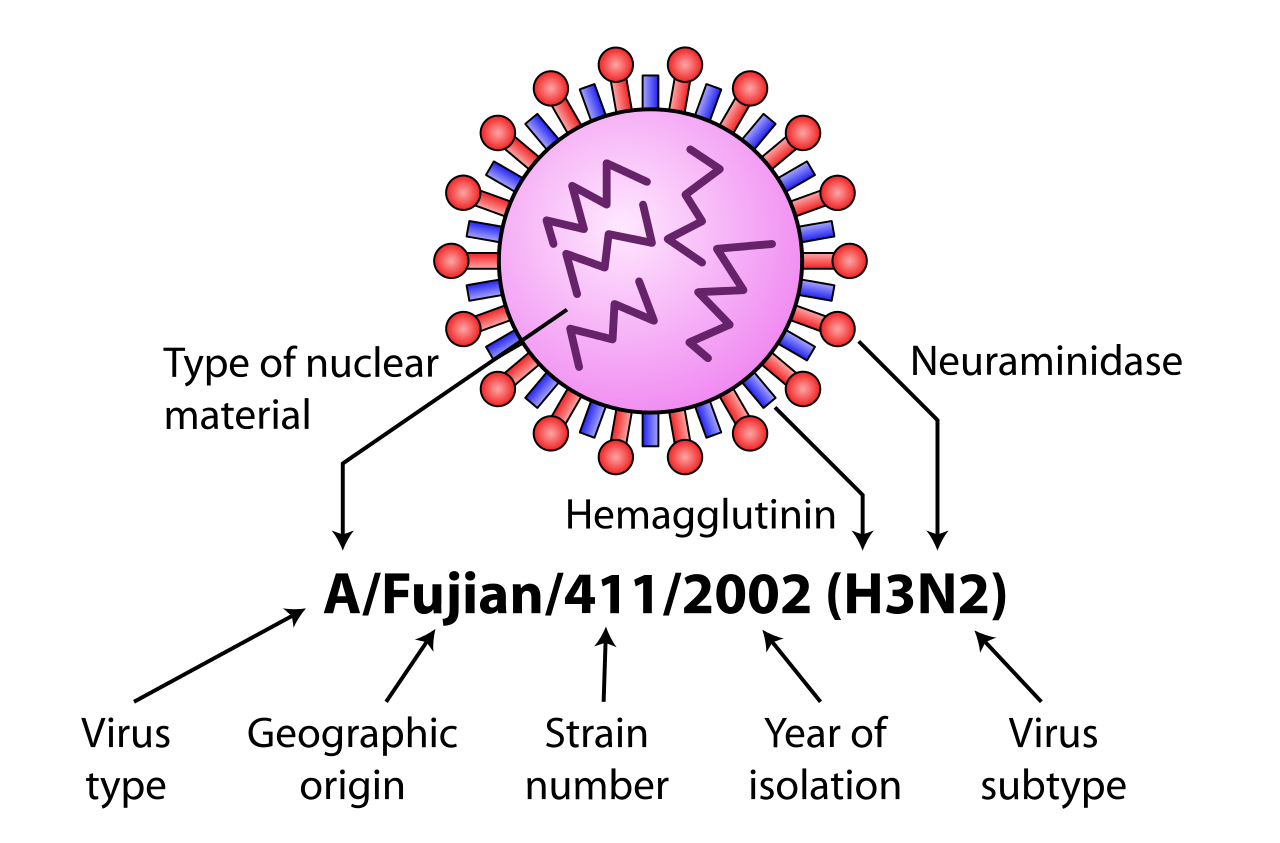
A型インフルエンザウイルスは、ウイルス表面タンパク質であるヘマグルチニン(HAまたはH)とノイラミニダーゼ(NAまたはN)に基づいてさらに分類されます。自然界では、18種類のHA亜型(血清型)と11種類のNA亜型が分離されています。これらのうち、HA亜型1~16とNA亜型1~9は野生の水鳥やシギ・チドリ類に見られ、HA亜型17~18とNA亜型10~11はコウモリからのみ分離されています。[28] [29]
さらに変異も存在し、特定のインフルエンザ株分離株はインフルエンザウイルス命名法[30]によって識別され、ウイルスの種類、宿主種(ヒト以外の場合)、最初に分離された地理的な場所、実験室の参照、分離年、HAおよびNAサブタイプが指定されます。[31] [32]
命名法の例は次のとおりです。
- A/ブリスベン/59/2007 (H1N1) - ヒトから分離
- A/豚/サウスダコタ/152B/2009 (H1N2) - 豚から分離
A型インフルエンザウイルスは、3種類のインフルエンザの中で最も毒性が強く、最も重篤な疾患を引き起こします。アウトブレイクやパンデミックを引き起こすA型インフルエンザウイルスはすべて、野生の水鳥に由来すると考えられています。[33] 1900年代以降のA型インフルエンザウイルスによるパンデミックはすべて、鳥インフルエンザが他のインフルエンザ株(ヒトに影響を及ぼす季節性インフルエンザ)または他の動物に影響を及ぼす株(2009年の豚インフルエンザパンデミックを参照)と遺伝子再集合によって引き起こしたものです。[34]ヒトで確認されている血清型は、死亡者数の多い順に以下のとおりです。
- H1N1は1918年に「スペイン風邪」、 2009年に「豚インフルエンザ」を引き起こした。 [35]
- H2N2は「アジア風邪」を引き起こした。
- H3N2は「香港風邪」を引き起こした。
- H5N1、「鳥インフルエンザ」。[36]
- H7N7は人獣共通感染の潜在性が異常である。[37]
- H1N2は豚と人間に感染します。[38]
- H9N2、H7N2、H7N3、H10N7。
| パンデミックの名前 | 日付 | 死亡者(数 | 致死率 | 関与するサブタイプ | パンデミック重症度指数 |
|---|---|---|---|---|---|
| 1889~1890年のインフルエンザパンデミック (アジア風邪またはロシア風邪)[41] |
1889–1890 | 100万 | 0.15% | H3N8 またはH2N2の可能性あり |
— |
| 1918年のスペイン風邪パンデミック (スペインかぜ)[42] |
1918~1920年 | 2000万から1億 | 2% | H1N1 | 5 |
| アジア風邪 | 1957–1958 | 100万から150万 | 0.13% | H2N2 | 2 |
| 香港風邪 | 1968~1969年 | 0.75から100万 | <0.1% | H3N2 | 2 |
| ロシア風邪 | 1977–1978 | 正確な数え方なし | — | H1N1 | — |
| 2009年のインフルエンザパンデミック[43] [44] | 2009~2010年 | 105,700~395,600 [45] | 0.03% | H1N1 | 該当なし |
インフルエンザB

インフルエンザBウイルスはほぼ完全にヒト病原体であり、インフルエンザAよりも一般的ではありません。インフルエンザBに感染しやすいことが知られている唯一の他の動物はアザラシです。 [ 46]このタイプのインフルエンザはA型よりも2〜3倍低い速度で変異し[47]、その結果遺伝的多様性が低く、インフルエンザBの血清型は1つだけです。[25]この抗原多様性の欠如の結果として、インフルエンザBに対するある程度の免疫は通常若い年齢で獲得されます。しかし、インフルエンザBは永続的な免疫が不可能なほどに変異します。[48]この抗原変化率の低下と、宿主域の制限(種間の抗原シフトを阻害する)が相まって、インフルエンザBのパンデミックは発生しません。[49]
インフルエンザC
C型インフルエンザウイルスはヒトや豚に感染し、重篤な疾患や地域的な流行を引き起こす可能性があります。[50]しかし、C型インフルエンザは他のタイプよりも一般的ではなく、通常は小児に軽度の疾患を引き起こします。[51] [52]
インフルエンザD
この属は2016年に分類され、そのメンバーは2011年に初めて分離されました。[53]この属はC型インフルエンザに最も近縁であると考えられており、数百年前にそこから分岐しました。[54]この属には少なくとも2つの株が現存しています。[55]主な宿主は牛と思われますが、このウイルスは豚にも感染することが知られています。
疫学
進化と歴史
.jpg/1280px-Genetic_Relationships_Among_Human_and_Swine_Influenza_Viruses,_1918-2009_(7704014350).jpg)
インフルエンザウイルスの主な自然宿主は野生の水鳥であると考えられている。[56] 2023年に行われた全RNAシーケンシングとトランスクリプトームマイニングの研究により、インフルエンザウイルスおよびより広義のArticulavirales目は水生起源である可能性が高く、魚類は最も初期の宿主の一部であり、この目の中で最も初期のウイルスは6億年以上前に甲殻類から進化したことが明らかになった。 [57] A型インフルエンザウイルスの亜型は2,000年前に分岐したと推定されている。A型インフルエンザウイルスとB型インフルエンザウイルスは約4,000年前に単一の祖先から分岐したと推定されているが、A型インフルエンザウイルスとB型インフルエンザウイルスの祖先とC型インフルエンザウイルスの祖先は約8,000年前に共通の祖先から分岐したと推定されている。[58]
インフルエンザ様疾患の発生は、記録に残る歴史を通して確認されています。最初の記録は、紀元前412年にヒポクラテスによって残されたものと考えられます。 [59]歴史家藤川は、862年から1868年の間に日本で発生したインフルエンザ様疾患の流行を46件記録しています。[60]ヨーロッパとアメリカ大陸では、中世から19世紀末にかけて、数多くの流行が記録されています。 [59]

1918年から1919年にかけて、20世紀最初のインフルエンザの大流行(通称「スペイン風邪」)が発生し、世界中で推定2,000万人から5,000万人の死者を出した。現在では、このパンデミックは免疫学的に新しいA型インフルエンザのH1N1亜型によって引き起こされたことが分かっている。[61]次のパンデミックは1957年に発生した「アジア風邪」で、これはH2N2亜型のウイルスによって引き起こされた。この亜型のウイルスでは、HAとNAをコードするゲノムセグメントが鳥インフルエンザ株から再集合によって派生したものとみられるが、ゲノムの残りの部分は1918年のウイルスの子孫である。[62] 1968年のパンデミック(「香港風邪」)はH3N2亜型によって引き起こされた。この亜型では、NAセグメントが1957年のウイルスに由来し、HAセグメントが鳥インフルエンザ株から再集合されたものであった。[62]
21世紀には、H1N1インフルエンザの株(以降「H1N1pdm09」と命名)が、以前のH1N1株とは抗原的に大きく異なり、2009年にパンデミックを引き起こしました。豚の間で循環しているいくつかの株と非常によく似ているため、「豚インフルエンザ」として知られるようになりました。[63]
A型インフルエンザウイルスは鳥類や豚の間で循環し、進化を続けています。野鳥からは、H(1~16)とN(1~11)のほぼすべての組み合わせが分離されています。[64] 2024年6月現在、野鳥の個体群では、特に毒性の強い2つのIAV株、H5N1とH7N9が優勢です。これらは家禽類で頻繁に発生し、家禽類と濃厚接触したヒトへの流出感染も稀に発生します。 [65] [66]
パンデミックの可能性
インフルエンザウイルスは、 RNAウイルスの特徴である比較的高い変異率を有する。[67] A型インフルエンザウイルスのゲノムの分節構造は、2つの異なるインフルエンザウイルス株に同時に感染した宿主において、分節再集合による遺伝子組換えを容易にする。 [68] [69]株間の再集合により、ヒトには影響を与えない鳥類株が、異なる株の特徴を獲得し、ヒトに感染してヒト間で伝播する、いわゆる人獣共通感染症が発生することがある。[70] 1900年代以降にヒトの間でアウトブレイクやパンデミックを引き起こしたA型インフルエンザウイルスはすべて、野生の水鳥に生息する株が他のインフルエンザ株と再集合して発生したものと考えられている。[71] [72]豚が再集合の中間宿主となる可能性もある(確実ではないが)。[73]
監視
世界インフルエンザ監視対応システム(GISRS)は、インフルエンザの蔓延を監視する世界的な研究所ネットワークであり、世界保健機関にインフルエンザの制御情報を提供し、ワクチン開発に役立てることを目的としています。[74] GISRSネットワークでは、127か国の研究所ネットワークを通じて、毎年数百万の検体が検査されています。[75] GISRSは、ヒトウイルスだけでなく、鳥、豚、その他の人獣共通感染性のインフルエンザウイルスも監視しています。
季節性インフルエンザ

インフルエンザシーズンは、インフルエンザA型またはインフルエンザB型によって引き起こされるインフルエンザの流行が毎年繰り返される時期です。このシーズンは、温帯地域では1年の寒い時期に発生し、北半球では11月から2月、南半球では5月から10月にかけて発生します。熱帯および亜熱帯地域でもインフルエンザシーズンが発生しますが、地域によって異なります。[77]毎年、世界中で季節性インフルエンザにより約300万~500万人が重症化し、29万人~65万人が死亡しています。[78]
温帯地域で冬にピークを迎える理由はいくつか考えられます。
- 冬は窓を閉め切った屋内で過ごす時間が長くなるため、インフルエンザ患者と同じ空気を吸い込み、ウイルスに感染する可能性が高くなります。[79]
- 冬は日照時間が短く、日光不足はビタミンDとメラトニンの分泌量の低下につながります。どちらも日光によって生成されます。その結果、免疫システムが弱まり、ウイルスに対する抵抗力が低下します。[79]
- インフルエンザウイルスは、より寒く乾燥した気候でよりよく生存し、したがってより多くの人々に感染することができる可能性がある。[79]
- 冷たい空気は鼻粘膜の感染抵抗力を低下させます。[80]
人獣共通感染症
人獣共通感染症は、細菌やウイルスなどの病原体がヒト以外の生物からヒトに感染することで起こるヒトの疾患である。[81] [82]鳥インフルエンザウイルスや豚インフルエンザウイルスは、稀にヒトに感染し、人獣共通インフルエンザウイルス感染症を引き起こすことがある。これらの感染症は通常、感染した動物や、感染した糞便や肉などの物質と密接に接触した人に限定され、他の人には感染しない。ヒトにおけるこれらの感染症の症状は非常に多様で、無症状または軽症の場合もあるが、重篤な疾患を引き起こし、重度の肺炎や死に至ることもある。[83]インフルエンザAウイルスの様々なサブタイプが人獣共通感染症を引き起こすことがわかっている。[83] [84]
人獣共通感染症は、良好な衛生状態を保つこと、家畜と野生動物の接触を防ぐこと、適切な個人用保護具を使用することで予防できます。[82]
2024年6月現在、世界中の野鳥の間で蔓延している鳥インフルエンザの2つの亜型、H5N1型とH7N9型が懸念されています。どちらも家禽類に壊滅的な被害を与える可能性があり、どちらもヒトに感染し、比較的高い致死率を示しています。[84]特にH5N1型は幅広い哺乳類に感染しており、哺乳類宿主に適応している可能性があります。[85]
兆候と症状
人間

季節性インフルエンザの症状は風邪と似ていますが、通常はより重症で、鼻水が出る可能性は低いです。[89]症状の発症は突然で、初期症状は主に非特異的です。突然の発熱、筋肉痛、咳、倦怠感、喉の痛み、頭痛、睡眠障害、食欲不振、下痢または腹痛、吐き気、嘔吐などです。[90]
人間はまれに鳥インフルエンザや豚インフルエンザに感染することがありますが、通常は感染した動物や汚染された物質との密接な接触が原因です。症状は一般的に季節性インフルエンザに似ていますが、まれに重篤化し、死に至ることもあります。[91] [92]
その他の動物
鳥
野生水鳥の中には、様々な種類のA型インフルエンザウイルスを自然発生的に無症候性キャリアとして保有する種があり、毎年の渡りの過程で広範囲にウイルスを拡散させる可能性があります。 [93]鳥インフルエンザの症状は、感染の原因となるウイルスの種類と、感染した鳥の種類によって異なります。鳥におけるインフルエンザの症状には、頭部の腫れ、涙目、反応の鈍化、協調運動障害、くしゃみやゴロゴロといった呼吸困難などがあります。[94]
高病原性鳥インフルエンザ
鳥インフルエンザは経済的に重要な養鶏場に多大な影響を及ぼすため、鳥ウイルス株は高病原性(したがって、強力な制御措置が必要となる可能性がある)または低病原性に分類されます。この分類は鶏への影響のみに基づいています。ウイルス株は、意図的に感染させた鶏の75%以上が死亡した場合、または遺伝的に類似している場合、高病原性鳥インフルエンザと分類されます。もう1つの分類は、低病原性鳥インフルエンザです。[95]ウイルス株を低病原性または高病原性株に分類することは、飼育鶏における症状の重症度に基づいており、他の種における症状の重症度を予測するものではありません。低病原性鳥インフルエンザに感染した鶏は軽度の症状または無症状ですが、高病原性鳥インフルエンザは重篤な呼吸困難、産卵数の大幅な減少、そして突然死を引き起こします。[96]
2006年以来、世界獣疫事務局は、低病原性鳥インフルエンザH5およびH7亜型が高病原性株に変異する可能性があるため、検出された場合はすべて報告することを義務付けている。[97]
豚
豚における豚インフルエンザの症状としては、発熱、抑うつ状態、咳(吠え)、鼻水や目からの分泌物、くしゃみ、呼吸困難、目の充血や炎症、食欲不振などが挙げられます。しかしながら、インフルエンザに感染した豚の中には、全く症状を示さないものもあります。豚インフルエンザの亜型は主にH1N1、H1N2、H3N2であり[98]、豚同士の濃厚接触、または農場間で汚染された機器を移動させることによって感染が広がります[99] 。豚と濃厚接触した人間も感染することがあります[100]。
馬
馬インフルエンザは、馬、ロバ、ラバに感染する可能性があります。[101]馬の間での感染率は非常に高く、潜伏期間は1~3日と比較的短いです。[102]馬インフルエンザの臨床症状には、発熱、鼻水、乾燥した空咳、抑うつ、食欲不振、衰弱などがあります。[102] EIは、鳥インフルエンザAウイルスから進化したインフルエンザAウイルスの2つの亜型、H7N7とH3N8によって引き起こされます。[103]
犬
犬インフルエンザAに感染した動物のほとんどは、咳、鼻水、発熱、無気力、目やに、食欲不振などの症状を示し、2~3週間続きます。[104]犬インフルエンザAウイルスには2種類あり、1つはH3N8型、もう1つはH3N2型です。[104] H3N8型は馬インフルエンザ鳥インフルエンザウイルスから進化したもので、犬間で持続的に感染するように適応しました。H3N2型は、2004年に韓国または中国で犬に感染した鳥インフルエンザウイルスに由来しています。[104]このウイルスは、動物保護施設や犬舎だけでなく、食肉生産のために犬を飼育する農場でも生息している可能性があります。[105]
コウモリ
最初のコウモリインフルエンザウイルスであるIAV(H17N10)は、2009年にグアテマラのヒメキバタコウモリ(Sturnira lilium)から初めて発見されました。[106] 2012年には、2番目のコウモリインフルエンザAウイルスIAV(H18N11)がペルーのヒラタコウモリ(Artibeus planirostris)から発見されました。[107] [108] [109]コウモリインフルエンザウイルスは、コウモリ以外の種には適応しにくいことがわかっています。[110]
生存性と消毒
哺乳類インフルエンザウイルスは不安定な傾向があるが、宿主の粘液中では数時間生存することができる。[111]鳥インフルエンザウイルスは、室温で蒸留水中で100日間、17℃(63°F)で200日間生存することができる。鳥インフルエンザウイルスは堆肥中ではより早く不活化されるが、ケージ内の糞便中では最大2週間生存することができる。鳥インフルエンザウイルスは冷凍保存すると無期限に生存することができる。[111]インフルエンザウイルスは、漂白剤、70%エタノール、アルデヒド、酸化剤、第四級アンモニウム化合物に弱い。133°F(56°C)で最低60分間の加熱、およびpH2未満の低pHによって不活化される。[111]
ワクチン接種と予防

インフルエンザウイルス感染症の予防および治療には、ワクチンと薬剤が利用可能です。ワクチンは、H1N1型およびH3N2型のヒトインフルエンザAウイルス、ならびにインフルエンザBウイルスの不活化または弱毒生ウイルス粒子から構成されています。野生型ウイルスの抗原性は変化するため、ワクチンは毎年、種株を更新することで処方変更されています。[112]
より具体的には、インフルエンザワクチンは遺伝子再集合法を用いて作られており、この方法は 50 年以上使用されてきました。この方法では、科学者は非感染性のインフルエンザ株 1 種と感染性の株 1 種の両方を鶏卵に注入します。不活性株は鶏卵で非常によく増殖するものでなければなりません。科学者は、最終製品が感染を防ぐべき HA 受容体および N 受容体を持つ感染性株を選びます。科学者は、公衆衛生上最も多く循環している表面 HA および NA 型、および次のインフルエンザシーズンに流行する可能性が最も高いと考えられる型を選ぶことで、これらの株を選びます。病原性と非病原性の 2 つの株は、不活性株が感染性株の 2 つの糖タンパク質標的のコピーを 8 つ持つまで、増殖して DNA を交換します。最終的に、新しく作成されたウイルスの中から、科学者は鶏卵で最もよく増殖し、必要な HA 遺伝子と NA 遺伝子も持つ 6 つのバージョンを選びます。最終的には、望ましいタンパク質を含む非感染性の株を何百万個もの卵子に注入し、遺伝子を採取してワクチン製品に使用します。[112]
ワクチンを製造するもう一つの方法は、感染株から遺伝子を組み換え、実験室で複製を作成することです。鶏卵培養という面倒なプロセスを必要とせず、ウイルスプラスミドを用いて標的遺伝子を抽出します。[112]
種子株と野生ウイルスの抗原性が一致しない場合、ワクチンはワクチンを防御することができません。[112]
インフルエンザ治療薬としては、M2プロトンチャネルに阻害することでウイルス粒子の脱殻を阻害するアマンタジンとリマンタジン、およびNAに阻害することで感染細胞からのウイルス粒子の放出を阻害するオセルタミビル(商品名タミフル)、ザナミビル、ペラミビルなどがある。しかし、前者ではエスケープ変異体の発生頻度が高く、後者では発生頻度は低い。[113]
参照

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
参考文献
- ^ 国際ウイルス分類委員会 ウイルス索引 — オルトミクソウイルス (2006). ICTVdB—ユニバーサルウイルスデータベース バージョン4. Büchen-Osmond, C (編), コロンビア大学, ニューヨーク.
- ^ Jones LD, Nuttall PA (1989). 「トゴトウイルスの非ウイルス血症伝播:時間と距離の影響」Trans. R. Soc. Trop. Med. Hyg . 83 (5): 712–14 . doi :10.1016/0035-9203(89)90405-7. PMID 2617637.
- ^ Ely B (1999). 「感染性サケ貧血」.ミルヒルエッセイ.国立医学研究所. 2007年8月24日時点のオリジナルよりアーカイブ。 2007年9月14日閲覧。
- ^ レイナード RS、マレー AG、グレゴリー A (2001)。 「スコットランドの野生魚における伝染性サケ貧血ウイルス」。ディス。アクアト。組織。46 (2): 93–100 .土井: 10.3354/dao046093。PMID 11678233。
- ^ Noda T (2012-01-03). 「インフルエンザウイルス粒子の自然形態」. Frontiers in Microbiology . 2 : 269. doi : 10.3389/fmicb.2011.00269 . PMC 3249889. PMID 22291683 .
- ^ Einav T, Gentles LE, Bloom JD (2020-07-23). 「SnapShot: 数字で見るインフルエンザ」(PDF) . Cell . 182 (2): 532–532.e1. doi :10.1016/j.cell.2020.05.004. PMID 32707094. S2CID 220715148.
- ^ ab 「ICTV第9回報告書; 2009年分類リリース:オルソミクソウイルス科」. ICTV . 2020年9月19日閲覧。
- ^ Ghedin E, Sengamalay NA, Shumway M, Zaborsky J, Feldblyum T, Subbu V, Spiro DJ, Sitz J, Koo H, Bolotov P, Dernovoy D, Tatusova T, Bao Y, St George K, Taylor J, Lipman DJ, Fraser CM, Taubenberger JK, Salzberg SL (2005年10月). 「ヒトインフルエンザの大規模シーケンシングは、ウイルスゲノム進化の動的な性質を明らかにする」. Nature . 437 (7062): 1162–6 . Bibcode :2005Natur.437.1162G. doi : 10.1038/nature04239 . PMID 16208317.
- ^ 鈴木雄三 (2005年3月). 「インフルエンザのシアロ生物学:インフルエンザウイルスの宿主域変異の分子メカニズム」. Biological & Pharmaceutical Bulletin . 28 (3): 399– 408. doi : 10.1248/bpb.28.399 . PMID 15744059.
- ^ abc Hilleman MR (2002年8月). 「ヒトウイルス性インフルエンザの実態と謎:病因、疫学、そして制御」.ワクチン. 20 ( 25–26 ): 3068–87 . doi :10.1016/S0264-410X(02)00254-2. PMID 12163258.
- ^ Wilson JC, von Itzstein M (2003年7月). 「新しい抗インフルエンザ療法の探索における最近の戦略」. Current Drug Targets . 4 (5): 389– 408. doi :10.2174/1389450033491019. PMID 12816348.
- ^ Bouvier NM, Palese P (2008年9月). 「インフルエンザウイルスの生物学」. Vaccine . 26 (Suppl 4): D49–53. doi :10.1016/j.vaccine.2008.07.039. PMC 3074182. PMID 19230160 .
- ^ Suarez DL, Spackman E, Senne DA, Bulaga L, Welsch AC, Froberg K (2003). 「リアルタイムRT-PCRによる鳥インフルエンザウイルス検出における各種消毒剤の効果」. Avian Diseases . 47 (3 Suppl): 1091–5 . doi :10.1637/0005-2086-47.s3.1091. PMID 14575118. S2CID 8612187.
- ^ 「鳥インフルエンザ(鳥インフルエンザ)のヒト疾患への影響。インフルエンザAウイルスの物理的特性」。CIDRAP - 感染症研究政策センター。ミネソタ大学。2024年3月12日。
- ^ 「インフルエンザウイルスは氷上で数十年生存可能」ニュージーランド・ヘラルド、ロイター通信、2006年11月30日。 2011年11月1日閲覧。
- ^ ab Wagner R, Matrosovich M, Klenk H (2002年5~6月). 「インフルエンザウイルス感染におけるヘマグルチニンとノイラミニダーゼの機能的バランス」Rev Med Virol . 12 (3): 159–66 . doi :10.1002/rmv.352. PMID 11987141. S2CID 30876482.
- ^ Lakadamyali M, Rust M, Babcock H, Zhuang X (2003年8月5日). 「インフルエンザウイルス感染の可視化」Proc Natl Acad Sci USA . 100 (16): 9280–85 . Bibcode :2003PNAS..100.9280L. doi : 10.1073/pnas.0832269100 . PMC 170909. PMID 12883000 .
- ^ Cros J, Palese P (2003年9月). 「ウイルスゲノムRNAの核内外への輸送:インフルエンザウイルス、トゴトウイルス、ボルナ病ウイルス」. Virus Res . 95 ( 1–2 ): 3–12 . doi :10.1016/S0168-1702(03)00159-X. PMID 12921991.
- ^ Kash J, Goodman A, Korth M, Katze M (2006年7月). 「インフルエンザウイルス感染における宿主細胞応答と翻訳制御のハイジャック」. Virus Res . 119 (1): 111–20 . doi :10.1016/j.virusres.2005.10.013. PMID 16630668.
- ^ Nayak D , Hui E, Barman S (2004年12月). 「インフルエンザウイルスの集合と出芽」. Virus Res . 106 (2): 147–65 . doi :10.1016/j.virusres.2004.08.012. PMC 7172797. PMID 15567494.
- ^ 「キャップ・スナッチング」ViralZone.expasy . 2014年9月11日閲覧。
- ^ Dias A, Bouvier D, Crépin T, McCarthy AA, Hart DJ, Baudin F, Cusack S, Ruigrok RW (2009年4月). 「インフルエンザウイルスポリメラーゼのキャップスナッチングエンドヌクレアーゼはPAサブユニットに存在する」. Nature . 458 (7240): 914–8 . Bibcode :2009Natur.458..914D. doi :10.1038/nature07745. PMID 19194459. S2CID 4421958.
- ^ Drake J (1993年5月1日). 「RNAウイルスにおける自然突然変異率」Proc Natl Acad Sci USA . 90 (9): 4171–5 . Bibcode :1993PNAS...90.4171D. doi : 10.1073/pnas.90.9.4171 . PMC 46468. PMID 8387212 .
- ^ 「ウイルス分類:2024年版」国際ウイルス分類委員会. 2025年3月19日閲覧。
- ^ ab Hay A, Gregory V, Douglas A, Lin Y (2001年12月29日). 「ヒトインフルエンザウイルスの進化」. Philos Trans R Soc Lond B Biol Sci . 356 (1416): 1861–70 . doi :10.1098/rstb.2001.0999. PMC 1088562. PMID 11779385 .
- ^ 「鳥インフルエンザ(Bird Flu)」. 米国疾病予防管理センター. 2007年9月15日閲覧。
- ^ Kumar, Binod; Asha, Kumari; Khanna, Madhu; Ronsard, Larance; Meseko, Clement Adebajo; Sanicas, Melvin (2018年4月). 「新興インフルエンザウイルスの脅威:現状と治療・制御の新たな展望」Archives of Virology . 163 (4): 831– 844. doi :10.1007/s00705-018-3708-y. ISSN 1432-8798. PMC 7087104. PMID 29322273 .
- ^ Alexander, Dennis J. (2007-07-26). 「鳥インフルエンザの疫学概観」. Vaccine . 25 (30): 5637– 5644. doi :10.1016/j.vaccine.2006.10.051. ISSN 0264-410X. PMID 17126960.
- ^ Wu, Ying; Wu, Yan; Tefsen, Boris; Shi, Yi; Gao, George F. (2014年4月). 「コウモリ由来インフルエンザ様ウイルスH17N10およびH18N11」. Trends in Microbiology . 22 (4): 183– 191. doi :10.1016/j.tim.2014.01.010. ISSN 0966-842X. PMC 7127364. PMID 24582528 .
- ^ 「インフルエンザウイルス命名法の改訂:WHO覚書」『世界保健機関誌』第58巻第4号、585~ 591頁、1980年。PMC 2395936。PMID 6969132。この覚書は 、
1980年2月にジュネーブで開催された会議の際に、590ページに記載されている署名者によって起草された
。 - ^ Atkinson W, Hamborsky J, McIntyre L, Wolfe S編 (2007). 『ワクチン予防可能疾患の疫学と予防』(第10版). ワシントンD.C.: 疾病管理予防センター.
- ^ 「鳥インフルエンザ:ヒト疾患への影響」ミネソタ大学感染症研究政策センター2007年6月27日2007年9月14日閲覧。
- ^ Webster, RG; Bean, WJ; Gorman, OT; Chambers, TM; Kawaoka, Y. (1992年3月). 「インフルエンザAウイルスの進化と生態」. Microbiological Reviews . 56 (1): 152– 179. doi :10.1128/mr.56.1.152-179.1992. ISSN 0146-0749. PMC 372859. PMID 1579108 .
- ^ Taubenberger, Jeffery K.; Morens, David M. (2010年4月). 「インフルエンザ:かつてのパンデミックと未来のパンデミック」.公衆衛生報告書. 125 (Suppl 3): 16– 26. doi :10.1177/00333549101250S305. ISSN 0033-3549. PMC 2862331. PMID 20568566 .
- ^ Wang TT, Palese P (2009年6月). 「豚インフルエンザウイルスの謎を解明する」. Cell . 137 (6): 983–85 . doi : 10.1016/j.cell.2009.05.032 . PMID 19524497.
- ^ Taubenberger, JK, Morens, DM (2009年4月). 「パンデミックインフルエンザ ― H5N1型ウイルスのリスク評価を含む」Rev. Sci. Tech. Off. Int. Epiz . 28 (1): 187– 202. doi :10.20506/rst.28.1.1879. PMC 2720801. PMID 19618626 .
- ^ Fouchier R、Schneeberger P、Rozendaal F、Brookman J、Kemink S、Munster V、Kuiken T、Rimmelzwaan G、Schutten M、Van Doornum G、Koch G、Bosman A、Koopmans M、Osterhaus A (2004)。 「鳥インフルエンザ A ウイルス (H7N7) はヒトの結膜炎と関連しており、急性呼吸窮迫症候群の致死例」。Proc Natl Acad Sci USA。101 (5): 1356–61。ビブコード:2004PNAS..101.1356F。土井:10.1073/pnas.0308352100。PMC 337057。PMID 14745020。
- ^ Malik-Peiris JS, Poon LL, Guan Y (2009年7月). 「ヒトにおける新規ブタ由来インフルエンザAウイルス(S-OIV)H1N1ウイルスの出現」. J Clin Virol . 45 (3): 169– 173. doi :10.1016/j.jcv.2009.06.006. PMC 4894826. PMID 19540800 .
- ^ Potter CW (2001年10月). 「インフルエンザの歴史」. Journal of Applied Microbiology . 91 (4): 572–9 . doi :10.1046/j.1365-2672.2001.01492.x. PMID 11576290. S2CID 26392163.
- ^ 「パンデミックインフルエンザについて知っておくべき10のこと」世界保健機関、2005年10月14日。2009年9月23日時点のオリジナルよりアーカイブ。2009年9月26日閲覧。
- ^ Valleron AJ, Cori A, Valtat S, Meurisse S, Carrat F, Boëlle PY (2010年5月). 「1889年のインフルエンザパンデミックの伝播性と地理的拡散」. Proc. Natl. Acad. Sci. USA . 107 (19): 8778–81 . Bibcode :2010PNAS..107.8778V. doi : 10.1073/pnas.1000886107 . PMC 2889325. PMID 20421481 .
- ^ Mills CE, Robins JM , Lipsitch M (2004年12月). 「1918年パンデミックインフルエンザの伝染性」. Nature . 432 (7019): 904–06 . Bibcode :2004Natur.432..904M. doi :10.1038/nature03063. PMC 7095078. PMID 15602562 .
- ^ Donaldson LJ, Rutter PD, Ellis BM, et al. (2009). 「イングランドにおけるパンデミックA/H1N1 2009インフルエンザの死亡率:公衆衛生監視研究」BMJ . 339 b5213. doi :10.1136/bmj.b5213. PMC 2791802. PMID 20007665 .
- ^ 「ECDC日報 – パンデミック(H1N1)2009 – 2010年1月18日」(PDF)欧州疾病予防管理センター2010年1月18日 オリジナル(PDF)より2010年1月22日時点のアーカイブ。 2010年1月18日閲覧。
- ^ ダーウッド FS、イウリアーノ AD、リード C、メルツァー MI、シェイ DK、チェン PY、バンダラナヤケ D、ブライマン RF、ブルックス WA、ビュシー P、フェイキン DR、ファウラー KB、ゴードン A、ヒエン NT、ホービー P、ファン QS、カッツ MA、クリシュナン A、ラル R、モンゴメリー JM、モルバック K、ピーボディ R、プレサニスAM、Razuri H、Steens A、Tinoco YO、Wallinga J、Yu H、Vong S、Bresee J、Widdowson MA (2012 年 9 月)。 「2009 年パンデミック インフルエンザ A H1N1 ウイルスの流行の最初の 12 か月に関連する推定世界死亡率: モデリング研究」。ランセット。感染症(投稿原稿)。12 (9): 687– 95. doi :10.1016/S1473-3099(12)70121-4. PMID 22738893.オリジナルより2024年4月9日時点の アーカイブ(PDF) – Zenodo経由。
- ^ Osterhaus AD, Rimmelzwaan GF, Martina BE, Bestebroer TM, Fouchier RA (2000年5月). 「アザラシにおけるインフルエンザBウイルス」. Science . 288 (5468): 1051–3 . Bibcode :2000Sci...288.1051O. doi :10.1126/science.288.5468.1051. PMID 10807575.
- ^ 信澤 栄治、佐藤 功(2006年4月). 「ヒトインフルエンザA型およびB型ウイルスの変異率の比較」. Journal of Virology . 80 (7): 3675–8 . doi :10.1128/JVI.80.7.3675-3678.2006. PMC 1440390. PMID 16537638 .
- ^ Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y (1992年3月). 「インフルエンザAウイルスの進化と生態」. Microbiological Reviews . 56 (1): 152– 79. doi :10.1128/MMBR.56.1.152-179.1992. PMC 372859. PMID 1579108 .
- ^ Zambon MC (1999年11月). 「インフルエンザの疫学と病因」. The Journal of Antimicrobial Chemotherapy . 44 Suppl B (Suppl B): 3– 9. doi : 10.1093/jac/44.suppl_2.3 . PMID 10877456. S2CID 15968981.
- ^ 松崎雄三、菅原健、水田健、土屋英二、村木雄三、本郷誠、鈴木秀、中村憲一 (2002). 「1996年と1998年に山形市で発生したインフルエンザCウイルスの抗原性および遺伝学的特徴」J Clin Microbiol . 40 (2): 422–29 . doi :10.1128/JCM.40.2.422-429.2002. PMC 153379. PMID 11825952 .
- ^ 松崎裕、勝島直、永井裕、庄司正、板垣隆、坂本正、北岡秀、水田和、西村裕 (2006年5月1日)。 「小児におけるインフルエンザCウイルス感染症の臨床的特徴」。J 感染障害193 (9): 1229–35 .土井: 10.1086/502973。PMID 16586359。
- ^ 片桐 誠・大泉 明・本間 正治 (1983年7月). 「児童養護施設におけるC型インフルエンザのアウトブレイク」J Infect Dis . 148 (1): 51– 56. doi :10.1093/infdis/148.1.51. PMID 6309999.
- ^ Hause BM, Ducatez M, Collin EA, Ran Z, Liu R, Sheng Z, Armien A, Kaplan B, Chakravarty S, Hoppe AD, Webby RJ, Simonson RR, Li F (2013年2月). 「2011年にオクラホマ州で分離された、ヒトインフルエンザC型ウイルスと遠縁の新規ブタインフルエンザウイルス」. PLOS Pathogens . 9 (2) e1003176. doi : 10.1371/journal.ppat.1003176 . PMC 3567177. PMID 23408893 .
- ^ Sheng Z, Ran Z, Wang D, Hoppe AD, Simonson R, Chakravarty S, Hause BM, Li F (2014年2月). 「豚由来の新規インフルエンザC様ウイルスのゲノム的および進化学的特徴づけ」Archives of Virology . 159 (2): 249–55 . doi :10.1007/s00705-013-1815-3. PMC 5714291. PMID 23942954 .
- ^ Collin EA, Sheng Z, Lang Y, Ma W, Hause BM, Li F (2015年1月). 「牛におけるインフルエンザDウイルスの2つの異なる遺伝的および抗原性系統の共循環」. Journal of Virology . 89 (2): 1036–42 . doi :10.1128/JVI.02718-14. PMC 4300623. PMID 25355894 .
- ^ Mahmoud SM、Alison M、Knobler SL編 (2005). 「1. インフルエンザの物語」.パンデミックインフルエンザの脅威:私たちは準備ができているか?ワークショップ概要. 米国医学研究所(米国)微生物の脅威に関するフォーラム. ワシントンD.C.:米国アカデミー出版.
- ^ Mary E. Petrone; Rhys Parry; Jonathon CO Mifsud; Kate Van Brussel; Ian Vorhees; Zoe T. Richards; Edward C. Holmes (2023年2月16日). 「インフルエンザウイルスとArticulavirales目が水生起源であることを示す証拠」プレプリント. BioRxiv . doi : 10.1101/2023.02.15.528772 . 2025年11月27日閲覧。
- ^ 鈴木雄三、根井正人(2002年4月)「インフルエンザウイルスヘマグルチニン遺伝子の起源と進化」分子生物学と進化誌19 ( 4). オックフォード・アカデミック: 501–509 . doi : 10.1093/oxfordjournals.molbev.a004105 . PMID 11919291.
- ^ ab 「インフルエンザの歴史」www.flu.com . 2024年6月20日閲覧。
- ^ 清水 功(1997年10月). 「インフルエンザ流行の歴史とインフルエンザウイルスの発見」.日本臨床. 日本臨床. 55 (10): 2505-2511 . PMID 9360364.
- ^ 「CDCアーカイブ:1918年のパンデミック(H1N1ウイルス)」米国疾病予防管理センター(CDC) . 2019年3月20日. 2024年6月20日閲覧。
- ^ Knobler SL、Mack A、Mahmoud A、Lemon SM他(米国医学研究所微生物脅威フォーラム)(2005年)「インフルエンザの物語」パンデミックインフルエンザの脅威:私たちは準備ができているか?ワークショップ概要。米国科学アカデミー出版局(米国) 。 2024年6月20日閲覧。
- ^ 「2009 H1N1パンデミック(H1N1pdm09ウイルス)」CU.S .米国疾病予防管理センター(CDC) . 2019年6月11日. 2024年6月21日閲覧。
- ^ 「FluGlobalNet – 鳥インフルエンザ」. science.vla.gov.uk . 2024年6月5日閲覧。
- ^ 「次のパンデミック:H5N1とH7N9インフルエンザ?」GAVIワクチンアライアンス。2021年3月26日。 2024年6月21日閲覧。
- ^ 「インフルエンザ(鳥インフルエンザおよびその他の人獣共通感染症)」世界保健機関2023年10月3日. 2024年6月21日閲覧。
- ^ サンフアン R、ネボット MR、キリコ N、マンスキー LM、ベルショー R (2010 年 10 月)。 「ウイルスの変異率」。ウイルス学ジャーナル。84 (19): 9733–9748。土井:10.1128/JVI.00694-10。PMC 2937809。PMID 20660197。
- ^ Kou Z, Lei FM, Yu J, Fan ZJ, Yin ZH, Jia CX, Xiong KJ, Sun YH, Zhang XW, Wu XM, Gao XB, Li TX (2005年12月). 「中国のスズメから分離された鳥インフルエンザH5N1ウイルスの新しい遺伝子型」. Journal of Virology . 79 (24): 15460– 15466. doi :10.1128/JVI.79.24.15460-15466.2005. PMC 1316012. PMID 16306617 .
- ^ 世界保健機関(WHO)世界インフルエンザプログラムサーベイランスネットワーク(2005年10月)「アジアにおけるH5N1型鳥インフルエンザウイルスの進化」新興感染症誌. 11 (10): 1515–1521 . doi :10.3201/eid1110.050644. PMC 3366754. PMID 16318689 . 図1は、アジア型H5N1ウイルスの様々な分離株におけるヘマグルチニン遺伝子の遺伝的関連性を図式化したものである。
- ^ 「動物と人の間での鳥インフルエンザウイルスの伝染」米国疾病予防管理センター(CDC) 2024年5月15日。 2024年6月10日閲覧。
- ^ Taubenberger JK, Morens DM (2010年4月). 「インフルエンザ:かつてのパンデミックと未来のパンデミック」.公衆衛生報告書. 125 (Suppl 3): 16– 26. doi :10.1177/00333549101250S305. PMC 2862331. PMID 20568566 .
- ^ Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y (1992年3月). 「インフルエンザAウイルスの進化と生態」. Microbiological Reviews . 56 (1): 152– 179. doi :10.1128/mr.56.1.152-179.1992. PMC 372859. PMID 1579108 .
- ^ 「ヒトと豚における豚インフルエンザに関するファクトシート」欧州疾病予防管理センター2017年6月15日. 2024年6月13日閲覧。
- ^ Lee K, Fang J (2013). 世界保健機関歴史辞典. Rowman & Littlefield. ISBN 9780810878587。
- ^ 「GISRS – 世界インフルエンザ監視・対応システム70周年」世界保健機関(WHO)2022年9月19日。 2024年6月13日閲覧。
- ^ CDC米国インフルエンザシーズン概要(毎週更新)「肺炎とインフルエンザ(P&I)死亡率監視」セクションを参照www.cdc.gov、2020年9月30日アクセス
- ^ Hirve S, Newman LP, Paget J, Azziz-Baumgartner E, Fitzner J, Bhat N, Vandemaele K, Zhang W (2016年4月27日). 「熱帯および亜熱帯におけるインフルエンザの季節性:ワクチン接種の時期は?」PLOS ONE . 11 (4) e0153003. Bibcode :2016PLoSO..1153003H. doi : 10.1371/journal.pone.0153003 . PMC 4847850 . PMID 27119988.
- ^ 「インフルエンザ(季節性)」世界保健機関. 2024年6月22日閲覧。
- ^ abc 「季節の理由:なぜインフルエンザは冬に流行するのか」ハーバード大学大学院芸術科学部の大学院生グループ「Science in the News」 2014年12月1日。 2024年6月21日閲覧。
- ^ LaMotte S (2022年12月6日). 「科学者たちはついに、冬に風邪やインフルエンザにかかりやすい理由を解明した」CNN . 2024年6月21日閲覧。
- ^ "zoonosis". Merriam-Webster.com Dictionary . Merriam-Webster . 2019年3月29日閲覧。
- ^ ab 「人獣共通感染症 - 重要な事実」世界保健機関2020年7月29日. 2024年6月24日閲覧。
- ^ ab 「人獣共通インフルエンザ - 2022年年次疫学報告書」www.ecdc.europa.eu 2023年5月23日2024年6月24日閲覧。
- ^ ab 「人獣共通感染症の可能性のある世界的なAIV」。国連食糧農業機関(FAO)。2020年7月29日。 2024年6月24日閲覧。
- ^ Plaza PI, Gamarra-Toledo V, Euguí JR, Lambertucci SA (2024年3月). 「世界における高病原性鳥インフルエンザA(H5N1)ウイルスの哺乳類感染パターンの最近の変化」. Emerging Infectious Diseases . 30 (3): 444– 452. doi :10.3201/eid3003.231098. PMC 10902543. PMID 38407173 .
- ^ 「インフルエンザの症状と診断」米国疾病予防管理センター(CDC) 2019年7月10日. 2019年12月27日時点のオリジナルよりアーカイブ。 2020年1月24日閲覧。
- ^ 「インフルエンザの症状と合併症」米国疾病予防管理センター(CDC). 2019年2月26日. 2020年8月1日時点のオリジナルよりアーカイブ。2019年7月6日閲覧。
- ^ Call SA, Vollenweider MA, Hornung CA, Simel DL, McKinney WP (2005年2月). 「この患者はインフルエンザに罹患しているか?」JAMA . 293 (8): 987– 997. doi :10.1001/jama.293.8.987. PMID 15728170.
- ^ 「風邪とインフルエンザ」米国疾病予防管理センター(CDC) 2022年9月29日. 2024年6月25日閲覧。
- ^ 「インフルエンザ」.英国国民保健サービス. 2023年8月9日. 2024年6月25日閲覧。
- ^ 「鳥インフルエンザ:ガイダンス、データ、分析」GOV.UK 2021年11月18日. 2024年5月9日閲覧。
- ^ 「ヒトにおける豚インフルエンザ」欧州疾病予防管理センター(ECDC)2017年9月20日。 2024年6月17日閲覧。
- ^ 「鳥インフルエンザ:家禽類やその他の飼育下の鳥における発見と報告方法」環境・食糧・農村地域省および動植物衛生庁2022年12月13日. 2024年5月6日閲覧。
- ^ 「鳥インフルエンザ」英国王立鳥類保護協会(RSPB) . 2024年6月25日閲覧。
- ^ Alexander DJ, Brown IH (2009年4月). 「高病原性鳥インフルエンザの歴史」Revue Scientifique et Technique . 28 (1): 19– 38. doi :10.20506/rst.28.1.1856. PMID 19618616.
- ^ 「鳥類における鳥インフルエンザ」米国疾病予防管理センター(CDC) 2022年6月14日。 2024年5月6日閲覧。
- ^ 「国家H5/H7鳥インフルエンザ監視計画」。米国農務省動植物検疫局。2013年10月。
- ^ 「ヒトと豚における豚インフルエンザに関するファクトシート」欧州疾病予防管理センター2017年6月15日. 2024年6月25日閲覧。
- ^ 「豚における豚インフルエンザ(豚インフルエンザ)に関する重要な事実」米国疾病予防管理センター(CDC) 2018年10月3日。 2024年6月25日閲覧。
- ^ 「2023年:豚インフルエンザの発生」世界保健機関2024年3月30日2024年6月25日閲覧。
- ^ 「馬インフルエンザ」。WOAH – 国際獣疫事務局。2024年6月25日閲覧。
- ^ ab 「馬インフルエンザ:馬の呼吸器疾患:メルク獣医マニュアル」www.merckvetmanual.com。2016年11月15日時点のオリジナルよりアーカイブ。 2016年12月4日閲覧。
- ^ 「馬インフルエンザ」.米国疾病予防管理センター(CDC) . 2023年5月5日. 2024年6月25日閲覧。
- ^ abc 「犬インフルエンザ(犬インフルエンザ)に関する重要な事実」米国疾病予防管理センター(CDC) 2023年8月29日。 2024年6月25日閲覧。
- ^ Wasik BR, Voorhees IE, Parrish CR (2021年1月). 「犬と猫のインフルエンザ」. Cold Spring Harbor Perspectives in Medicine . 11 (1) a038562. doi :10.1101/cshperspect.a038562. PMC 7778219. PMID 31871238 .
- ^ 「コウモリインフルエンザ(インフルエンザ)」cdc.gov 2024年7月10日. 2025年3月23日閲覧。
- ^ 「コウモリインフルエンザウイルスの特徴」uniklinik-freiburg.de . 2020年6月30日閲覧。
- ^ 「コウモリから新たなインフルエンザウイルスが発見される」Nature . 503 (7475): 169. 2013. doi :10.1038/503169e . 2020年6月30日閲覧。
- ^ Ciminski K, Pfaff F, Beer M, Schwemmle M (2020年4月). 「コウモリが明らかにするインフルエンザAウイルスの適応力の真の力」. PLOS Pathogens . 16 (4) e1008384. doi : 10.1371/journal.ppat.1008384 . PMC 7161946. PMID 32298389 .
- ^ Ciminski K, Ran W, Gorka M, Lee J, Malmlov A, Schinköthe J, Eckley M, Murrieta RA, Aboellail TA, Campbell CL, Ebel GD, Ma J, Pohlmann A, Franzke K, Ulrich R, Hoffmann D, García-Sastre A, Ma W, Schountz T, Beer M, Schwemmle M (2019 年 12 月)。 「コウモリインフルエンザウイルスはコウモリの間で伝染しますが、コウモリ以外の種にはほとんど適応しません。」自然微生物学。4 (12): 2298–2309。土井:10.1038/s41564-019-0556-9。PMC 7758811。PMID 31527796。S2CID 202580293 。
- ^ abc Spickler AR (2016年2月). 「インフルエンザ」(PDF) .食料安全保障・公衆衛生センター. アイオワ州立大学. p. 7.
- ^ abcd Hood, Ernie (2006). 「インフルエンザワクチン製造に弾み」. Environmental Health Perspectives . 114 (2): A108 – A111 . doi :10.1289/ehp.114-a108. PMC 1367863. PMID 16451835 .
- ^ 鈴木雄三 (2006年10月). 「インフルエンザウイルスゲノムにおける自然選択」. Molecular Biology and Evolution . 23 (10). Oxford Academic: 1902–11 . doi : 10.1093/molbev/msl050 . PMID 16818477. 2022年1月21日時点のオリジナルよりアーカイブ。
さらに読む
- ホイル, L. (1969). インフルエンザウイルス. ウイルス学モノグラフ. 第4巻. シュプリンガー・フェアラーク. ISBN 978-3-211-80892-4. ISSN 0083-6591. OCLC 4053391.
外部リンク
{kind=link}
- Health-EU ポータル: EU はインフルエンザに対する世界的な対応の準備に取り組んでいます。
- インフルエンザ研究データベース: インフルエンザのゲノム配列と関連情報のデータベース。
- 欧州委員会—公衆衛生:パンデミック(H1N1)2009に関するEUの調整
- EMデータバンク(EMDB)の3Dインフルエンザウイルス関連構造
- ウイルスゾーン:オルトミクソウイルス科
