{kind=link}
{kind=link}

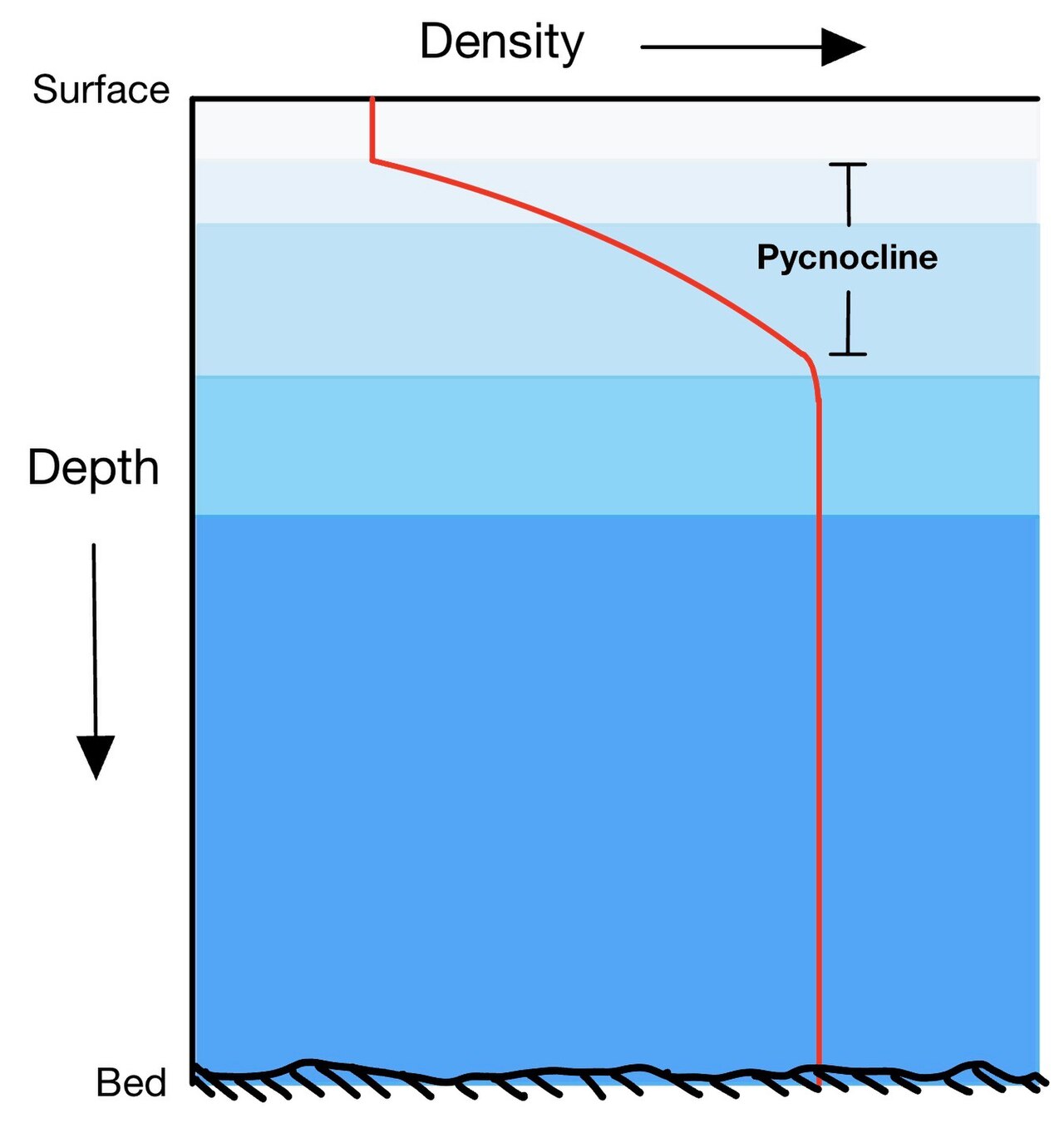
密度躍層とは、密度 勾配(∂ρ/∂z)は水域内で最大となる。海流は、砕波、温度・塩分濃度の差、風、コリオリの力、天体の重力による潮汐などの力によって発生する。さらに、密度勾配によって駆動される密度躍層の物理的特性も、海流や鉛直プロファイルに影響を与える。これらの変化は、海洋を通じた熱、塩分、栄養素の輸送と関連しており、密度躍層の拡散は湧昇を制御する。[1]
混合層の下では、安定した密度勾配(またはピクノクライン)が上層水と下層水を分離し、鉛直輸送を妨げています。[2]この分離は、海洋と海洋生物に重要な生物学的影響を及ぼします。しかし、ピクノクラインを横切る鉛直混合は海洋では一般的な現象であり、せん断力によって生じる乱流によって発生します。[3]このような混合は、栄養素の輸送において重要な役割を果たします。[4]
身体機能
風と波によって生じる乱流混合により、表層から下方へ熱が伝達されます。低・中緯度地域では、この混合により、深さ数メートルから数百メートルにわたって、ほぼ均一な温度の水の表層混合層が形成されます。この混合層の下、外洋では水深200~300メートルで、水温は1000メートル付近まで急激に低下します。温度勾配が最も急な水層は、永久サーモクラインと呼ばれます。[5]この層の温度差は、緯度によっては20℃にもなります。永久サーモクラインは、暖かく密度の低い表層水とその下にある冷たく密度の高い底層水との間の水密度の変化と一致しています。密度が急激に変化する領域は密度躍層と呼ばれ、鉛直方向の水循環の障壁として機能します。そのため、海の生物学で役割を果たす特定の化学物質の鉛直分布にも影響を与えます。温度と密度の急激な勾配は、動物の垂直方向の移動を制限する要因となる可能性もある。[6]
季節性
上記で説明した密度躍層の一般的な構造は当てはまりますが、密度躍層は季節によって変化することがあります。冬季は海面水温が低く、波が大きくなる傾向があるため、混合層の深さがメインのサーモクライン/密度躍層に至るまで増加する場合があります。[7]
夏には、気温の上昇、海氷と陸氷の融解、そして日照時間の増加により、海洋表層の温度が上昇します。この表層は、以前に形成された大きな冬季混合層の上に位置し、主密度躍層の上に季節性密度躍層を形成します。冬季混合層は密度勾配が小さくなり、密度勾配はピクノスタッドと呼ばれます。季節が再び変わり始めると、表層からの熱の純損失と継続的な風による混合により、季節性密度躍層は次の夏まで侵食されます。[7]
緯度による変化
温度と塩分濃度はどちらも密度に影響を与えますが、緯度地域によってはどちらかの影響が大きくなることがあります。熱帯および中緯度地域では、すべての海洋の表面密度は表面塩分濃度ではなく表面温度に従います。50度を超える高緯度地域では、温度が常に氷点付近にあるため、すべての海洋の表面密度は温度よりも塩分濃度に従います。[7]
低緯度および中緯度では、水深200~1000メートルに恒久的な密度躍層が存在します。大西洋のサルガッソ海のように、広大だが地理的に限定された亜熱帯地域では、2つの恒久的な温度躍層が存在し、サーモスタッドと呼ばれる下層の鉛直成層によって隔てられています。この現象は密度に反映され、密度は海水温に強く依存します。2つの恒久的な密度躍層は恒久的な温度躍層と関連しており、サーモスタッドに相当する密度は密度躍層と呼ばれます。[7]
亜極地および極地では、表層水は緯度の影響で年間を通して非常に冷たく、海氷と陸氷の融解、多雨、淡水流出の影響で非常に淡水化しています。一方、深層水は地球全体でほぼ一定です。このため、恒久的なサーモクライン(水温躍層)は存在しませんが、季節的なサーモクライン(水温躍層)は発生する可能性があります。これらの地域では恒久的なハロクライン(塩分躍層)が存在し、このハロクラインが恒久的なピクノクラインを決定する主な要因となっています。[7]
生物学的機能
{kind=link}
{kind=link}

植物プランクトンの成長速度は栄養塩濃度によって制御されており、海中における栄養塩の再生は、高栄養段階と低栄養段階の相互作用において非常に重要な役割を果たしている。ピクノクラインの形成による分離は、下層から上層への栄養塩の供給を阻害する。ピクノクラインを通過する栄養塩のフラックスは、他の表層よりも低い。[8]
微生物ループ
微生物ループは、海洋微生物食物網における栄養経路です。「微生物ループ」という用語は、Azamら(1983)によって造語され、海洋生態系の炭素・栄養循環において微生物が果たす役割を説明するために用いられました。この循環において、溶存有機炭素(DOC)は細菌バイオマスに取り込まれ、より高次の栄養段階へと戻されます。また、植物プランクトン、動物プランクトン、ネクトンによって形成される古典的な食物連鎖と結合しています。
植物プランクトンブルームの終焉、つまり藻類が老化期に入ると、植物デトリタスが蓄積し、溶解代謝物の放出が増加する。特にこの時期に、バクテリアはこれらのエネルギー源を利用して増殖し、植物プランクトンのブルームに続く急激なパルス(ブルーム)を引き起こす。植物プランクトンとバクテリアの同様の関係は、バクテリアプランクトンの垂直分布にも影響を与える。バクテリアの最大数は、通常、上部の有光層から沈降して植物デトリタスが蓄積するピクノクラインで発生する。そこでバクテリアによる分解は、安定した水域における酸素極小層の形成に寄与する。[9]
日周垂直移動
プランクトンの最も特徴的な行動の一つは、24時間周期で起こる垂直移動である。これはしばしば日周垂直移動または日周移動と呼ばれてきた。24時間で移動する垂直距離は様々で、一般的に大型の種や優れた遊泳力を持つ種の方が長い。しかし、小さなカイアシ類でさえ24時間以内に数百メートルを2回移動することもあり、オキアミ類やエビのようなより優れた遊泳力を持つものは800メートル以上移動することもある。[10]移動の深度範囲は、サーモクラインまたはピクノクラインの存在によって制限される可能性がある。しかし、日周垂直移動が可能な植物プランクトンと動物プランクトンは、多くの場合、ピクノクラインに集中している。[11]さらに、サーモクラインまたはピクノクラインを遊泳する能力を持つ海洋生物は、移動中に強い温度勾配と密度勾配、およびかなりの圧力変化を経験する可能性がある。
安定性
ピクノクラインは、リチャードソン数が0.25を下回ると不安定になります。リチャードソン数は、位置エネルギーと運動エネルギーの比を表す無次元値です。この比は、せん断速度が成層速度を超えると0.25を下回ります。これによりケルビン・ヘルムホルツ不安定性が生じ、乱流が発生して混合が促進されます。[12]
ピクノクラインの深度や特性の変化は、いくつかのコンピュータプログラムモデルでシミュレートできます。これらのモデルに対する最も簡単な方法は、海洋大循環モデル(OCGM)に基づくエクマンポンプモデルを調べることです。[13]
斜面の種類
参照
参考文献
- ^ アナンド・グナナデシカン. 1999. 海洋密度躍層の構造を予測するシンプルなモデル. Science 283 (5410): 2077–2079.
- ^ Mann and Lazier (2006). 海洋生態系のダイナミクス. 第3版. Blackwell Publishing. 第3章.
- ^ 成層流体における乱流混合、流体力学年次レビュー(1991年)
- ^ 成層せん断層を通じた鉛直混合と輸送、Journal of Physical Oceanography (2001)
- ^ クナウス、ジョン・A. (1997). 物理海洋学入門. 第2版, プレンティス・ホール. 第1章
- ^ Lalli and Parson (1993). 生物海洋学入門. ペルガモン出版. 第2章.
- ^ abcde タリー, リン・D.; ピカード, ジョージ・L.; エメリー, ウィリアム・J.; スウィフト, ジェームズ・H. (2011). 『記述的物理海洋学入門』(第6版). エルサーヴィエ. ISBN 978-0-7506-4552-2。
- ^ Hales, B., Hebert, D., Marra, J. 2009. ニューイングランド棚氷前線における植物プランクトンへの栄養塩の乱流供給. Journal of Geophysical Research. Vol. 114, C05010, doi :10.1029/2008JC005011.
- ^ Lalli and Parson (1993). 生物海洋学入門. ペルガモン出版. 第5章.
- ^ Lalli and Parson (1993). 生物海洋学入門. ペルガモン出版. 第4章.
- ^ Hill, AE 1998. 成層潮汐流における日周鉛直移動:プランクトン分散への影響. Journal of Marine Research , Vol 56, pp. 1069-1096.
- ^ 密度成層、乱流、しかし混合はどの程度か?流体力学年次レビュー(2008年)
- ^ Capotondi, A., Alexander, MA, Deser, C., Miller, A. 2004. 北東太平洋における低周波密度躍層変動. アメリカ気象学会誌. 第35巻, pp. 1403-1420.