昆虫の羽

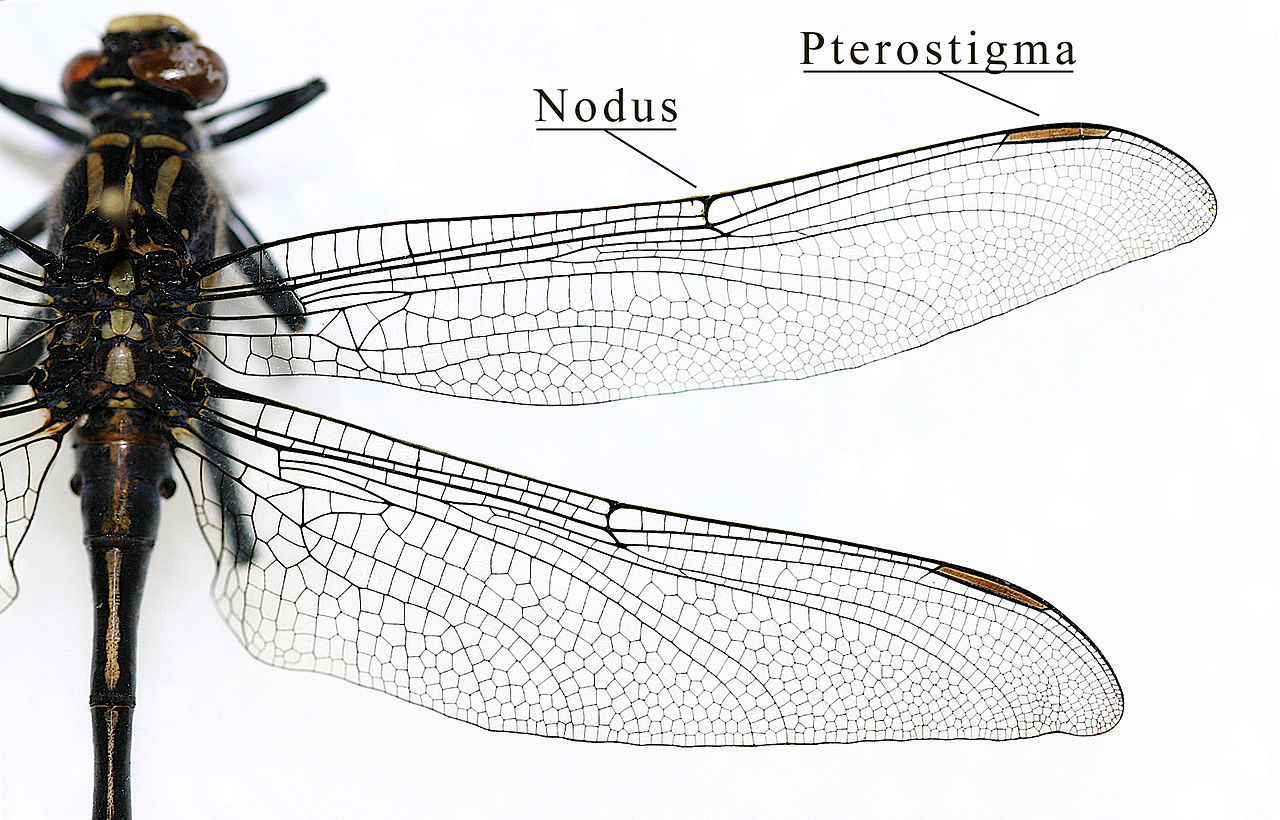



昆虫の翅は、昆虫の成虫が飛翔することを可能にする外骨格の発達部分である。第2胸節と第3胸節(中胸部と中胸部)に存在し、それぞれ前翅と後翅と呼ばれることが多いが、後翅、さらには原基さえも持たない昆虫もいる。翅は多数の縦脈によって強化されており、これらの縦脈はしばしば交差結合して膜状の閉鎖「細胞」を形成する(極端な例としてトンボやクサカゲロウが挙げられる)。翅脈の融合と交差結合によって生じる模様は、異なる進化系統を特徴づける特徴となることが多く、多くの昆虫目において科レベル、さらには属レベルまでの同定に用いることができる。
物理的に見ると、昆虫の中には飛翔筋を直接動かすものもあれば、間接的に動かすものもあります。直接飛翔する昆虫では、翅の筋肉が翅の基部に直接付着しているため、翅の基部をわずかに下方に動かすと、翅自体が上方に持ち上げられます。間接飛翔する昆虫では、胸部に付着して変形させる筋肉があり、これによって翅も動きます。
羽は、ビロードアントやストレプシプテラなどの一部のグループでは一方の性別(多くの場合はオス)にのみ存在し、アリやシロアリなどの社会性昆虫の「働きアリ」では選択的に失われている。まれに、イチジクバチのようにメスには羽がありオスにはないこともある。アブラムシの分散期など、ライフサイクルの特定の時期にのみ羽が生成される場合もある。羽の構造と色は、アブラムシ、イナゴの渡り期、多形性の蝶など、形態によって変化することが多い。静止時には、羽は平らに保たれているか、特定のパターンに沿って複数回折りたたまれている。最も一般的には後羽が折りたたまれているが、スズメバチなどのいくつかのグループでは前羽が折りたたまれている。
昆虫の羽の進化的起源については議論が続いている。19世紀、昆虫の羽の進化に関する問題は、当初2つの主要な立場に分かれていた。1つは、昆虫の羽は既存の構造から進化したという説であり、もう1つは、昆虫の羽は全く新しい構造であるという説であった。[ 1 ] [ 2 ]この「新しい」仮説は、昆虫の羽は既存の祖先の付属器からではなく、昆虫の体壁からの突起として形成されたと示唆した。[ 3 ]
昆虫の羽の起源に関する研究は、19世紀に提唱された「既存の構造」という立場に基づいて長年進められてきました。[ 2 ]近年の文献では、昆虫の羽の起源において重要な祖先構造がいくつか指摘されています。例えば、鰓、脚の呼吸付属器、胸郭の側方(傍胸背)および後外側への突出などが挙げられます。[ 4 ]
より最近の文献によると、候補としては鰓様構造、傍胸葉、甲殻類の背板などが挙げられます。後者は、昆虫が甲殻類を直接の祖先とする汎甲殻類節足動物であり、四肢発達の遺伝的メカニズムを共有していることを示唆する最近の昆虫遺伝学研究に基づいています。[ 3 ] [ 5 ] [ 6 ] [ 7 ] [ 8 ]
昆虫の羽の起源に関する他の説としては、傍胸背板説、鰓説、そして昆虫の羽の進化に関する二重説などがある。これらの説は、羽が傍胸背板(胸背板 )の延長である傍胸背板から発達したとする説[ 5 ] 、カゲロウ類の水生幼虫に見られる可動式の腹部鰓の変形であるとする説[ 5 ]、あるいは、昆虫の羽は、それぞれ既存の関節と気管構造を持つ既存の内節構造と外節構造の融合から生じたとする説[ 9 ] [ 10 ]を唱えている。
形態学

内部
それぞれの羽は、静脈系に支えられた薄い膜で構成されている。膜は2層の外皮が密接に接して形成され、静脈は2層が分離した場所に形成される。下側のクチクラは、静脈の下でより厚く、より強く硬化している場合もある。主要な静脈の内側には神経と気管があり、静脈の空洞は血体腔とつながっているため、血リンパが羽に流入することができる。[ 11 ]
翅が発達するにつれて、背側と腹側の外皮層は、そのほとんどの領域で密着し、翅膜を形成する。残りの領域は、将来静脈となる通路を形成し、神経や気管が形成される可能性がある。静脈を囲むクチクラは厚くなり、より強く硬化することで、翅に強度と剛性を与える。翅には2種類の毛が生える。小毛は小さく不規則に散在し、大毛はより大きく、ソケット状で、静脈のみに生える場合がある。鱗翅目と毛翅目の鱗粉は、大毛が大きく変化したものである。[ 12 ]
静脈

非常に小さな昆虫の中には、脈が大幅に減少しているものもある。例えば、ハチ目バチでは、肋下葉と橈骨の一部しか存在しない。逆に、既存の脈が分岐して副脈を形成したり、バッタ目(バッタやコオロギ)の羽のように、元の脈の間に介在脈が発達したりすることで、脈が増加することもある。昆虫の中には多数の横脈が存在するものもあり、トンボ目(トンボやイトトンボ)の羽や、キリギリス目(キリギリス科)とバッタ目(キリギリス科)の前羽の基部のように、横脈が網状構造を形成することもある。[ 11 ]
アーケディクティオンとは、最初の有翅昆虫について提唱された翅脈の仮説的な構造に付けられた名称である。これは推測と化石データの組み合わせに基づいている。すべての有翅昆虫は共通の祖先から進化したと考えられているため、アーケディクティオンは2億年にわたる自然淘汰によって修正(そして合理化)されてきた「鋳型」を表している。現在の仮説によれば、アーケディクティオンには6~8本の縦脈が含まれていたとされる。これらの脈(とその枝)は、ジョン・コムストックとジョージ・ニーダムによって考案されたコムストック・ニーダム方式に従って命名されている。[ 13 ]
- コスタ(C) – 翼の前縁
- 肋下静脈(Sc) - 肋骨の後ろにある第2縦脈。通常は分岐しない。
- 橈骨(R) - 第3縦脈、1~5本の枝が翼縁に達する
- 中脈(M) - 第4縦脈、1~4本の枝が翼縁に達する
- 肘(Cu) - 第5縦脈、1~3本の枝が翼縁に達する
- 肛門静脈(A1、A2、A3) – 肘の後ろにある分岐のない静脈
肋骨 (C) はほとんどの昆虫の主要な縁静脈である。肋骨の上に前肋骨と呼ばれる小さな静脈があることもあるが、現存するほぼすべての昆虫で[ 14 ] : 41–42 前肋骨は肋骨と癒合している。肋骨は前縁にあり、その基部は上腕骨板と関連しているため、分岐することはめったにない。肋静脈の気管はおそらく肋下気管の枝である。肋骨の後には 3 番目の静脈である肋下静脈があり、前静脈と後静脈の 2 つの別々の静脈に分岐する。肋下静脈の基部は、第 1 腋窩頸部の遠位端と関連している (以下のセクションを参照)。4 番目の静脈は橈骨 (R) で、5 つの別々の静脈に分岐する。橈骨静脈は一般に翅で最も強い静脈である。翼の中央に向かって、橈骨は第一の未分岐枝(R1)と第二の枝(橈骨扇状枝(Ra))に分岐し、橈骨扇状枝はさらに4つの遠位枝(R2、R3、R4、R5)に二分する。基底部では、橈骨は第二腋窩枝(2Ax)の前端と柔軟に結合している。[ 15 ]
翅の第5脈は中脈である。原型パターン(A)では、中脈は2つの主要な枝に分岐する。1つは前中脈(MA)で、これは2つの遠位枝(MA1、MA2)に分岐する。もう1つは、4つの末端枝(M1、M2、M3、M4)を持つ中脈、または後中脈(MP)である。ほとんどの現生昆虫では前中脈は失われており、通常の「中脈」は共通の基底茎を持つ4枝の後中脈である。カゲロウ目(Ephemeridae)では、現在の翅脈の解釈によれば、中脈の2つの枝は保持されているが、トンボ目(Tdonata)では残存する中脈は原始的な前枝である。中脈の茎は橈骨静脈と一体化していることが多いが、独立した静脈として存在する場合は、その基部が遠位正中板(m')と関連しているか、後者と連続的に硬化している。翅の第6脈である肘静脈は、主に2枝に分岐している。肘枝の主分岐は翼基部付近で起こり、2つの主枝(Cu1、Cu2)を形成する。前枝は複数の二次枝に分岐することもあるが、通常は2つの遠位枝に分岐する。膜翅目、トビ翅目、チョウ目における肘枝の第二枝(Cu2)は、コムストックとニーダムによって第一肛門枝と誤認された。肘枝の近位部では、主茎は翼基部の遠位正中板(m')と関連している。[ 15 ]
後肘 (Pcu) は、コムストック・ニーダム体系における最初の肛門である。しかし、後肘は独立した翅脈の地位を有し、そのように認識されるべきである。幼虫の翅では、後肘の気管は肘気管と前翅気管群の間から発生する。より一般的な昆虫の成熟した翅では、後肘は常に肘の近位に付随し、翅基部の屈筋小板 (3Ax) と密接に連結することは決してない。アオムシ目、メコプター目、トビフテリ目では、後肘は前翅脈とより密接に付随することがあるが、その基部は常に前翅脈から独立している。後肘は通常は分岐しないが、原始的には2分岐している。羽端静脈 (IV~NV) は、第 3 腋窩静脈に直接関連し、翅の屈曲を引き起こすこの小板の動きによって直接影響を受ける肛門静脈です。羽端静脈の数は、翅の羽端領域の拡張に応じて 1 本から 12 本まで変化します。羽端気管は通常、幼虫の昆虫では共通の気管茎から発生し、静脈は 1 本の肛門静脈の枝とみなされます。遠位では、羽端静脈は単純または分岐しています。翅の頬葉の頬静脈 (J) は、不規則な静脈のネットワークで占められていることが多く、完全に膜状の場合もありますが、第 1 頬静脈 (弓状静脈) と第 2 頬静脈 (心静脈) (2J) の 1 つまたは 2 つの明確な小静脈が含まれる場合もあります。[ 15 ]
- C-Sc横静脈– 肋骨と肋骨下の間を走る
- R横脈– 橈骨の隣接する枝の間を走る
- 橈骨横静脈– 橈骨と中膜の間を走る
- M-Cu横脈– 中膜と肘部の間を走る
翅のすべての脈は二次分岐し、交差脈によって合流する。昆虫のいくつかの目では、交差脈の数が非常に多いため、全体の脈脈パターンは分岐脈と交差脈の密接なネットワークとなる。しかしながら、通常は特定の位置に決まった数の交差脈が存在する。より一定数の交差脈は、肋骨と肋下骨の間の上腕横脈(h)、RとRsの最初の分岐の間の放射横脈(r)、R8の2つの分岐の間の扇形横脈(s)、M2とM3の間の正中横脈(m-m)、そして中腕と肘の間の中肘横脈(m-cu)である。[ 15 ]
昆虫の羽の静脈は、カゲロウに見られるような凸凹の配置(つまり、凹が下、凸が上)と、規則的に交互に現れる三角状の分岐を特徴とします。静脈が分岐するたびに、必ず2つの分岐の間には反対側の位置に挿入された静脈が存在します。凹状の静脈は2つの凹状の静脈に分岐し(挿入された静脈は凸状)、静脈の規則的な変化は維持されます。[ 16 ]羽の静脈は、羽を弛緩させたときに上向きに折りたたまれるか、下向きに折りたたまれるかによって、波状のパターンを形成します。静脈の基底部は凸状ですが、各静脈は遠位方向に分岐して前方凸状枝と後方凹状枝に分岐します。このように、肋骨と肋下骨は主第一脈の凸状と凹状の枝とみなされ、Rsは橈骨の凹状枝、後中膜は中膜の凹状枝、Cu1とCu2はそれぞれ凸状と凹状であり、原始的な後肘と第一前橈骨はそれぞれ前凸状枝と後凹状枝を持つ。脈の凸状または凹状の性質は、現生昆虫の脈の末端枝が残存しているかどうかを決定する証拠として用いられてきたが、すべての翅において一貫していることは証明されていない。[ 11 ] [ 15 ]
フィールド

翅の領域は、翅が折り畳まれる襞線と、飛行中に翅が曲がることができる屈曲線によって区切られ、さらに分割される。屈曲線と襞線の基本的な区別は、襞線がある程度柔軟性を許容する場合もあれば、その逆もあるため、しばしば曖昧である。ほぼ全ての昆虫の翅に見られる2つの不変の襞は、鎖骨襞(屈曲線)と頬襞(または襞線)であり、これらは可変的で不十分な境界を形成する。翅の折り畳みは非常に複雑で、皮膚翅目や鞘翅目の後翅では横方向の折り畳みが見られ、一部の昆虫では肛門領域が扇状に折り畳まれる。[ 14 ] 昆虫の翅には約4つの異なる領域が見られる。
- レミギウム
- 肛門部(ヴァンヌス)
- 頬部
- 腋窩部
- アルーラ
静脈と横静脈のほとんどは、胸筋の力で飛ぶ動作の大部分を担うレミギウムの前部にあります。レミギウムの後部は、時に楔状静脈と呼ばれます。他の 2 つの後部領域は、肛門領域と頬部領域です。[ 14 ]ヴァンナル ヒダが肛門静脈群の前方にある通常の位置にある場合、レミギウムには肋静脈、肋下静脈、橈静脈、内側静脈、肘静脈、肘後静脈が含まれます。翼を屈曲させた状態では、レミギウムは橈静脈と第二腋窩静脈の柔軟な基底部接続部で後方に曲がり、内側肘部領域の基底部は、翼基部の正中板 (m、m') の間にある基底襞 (bf) に沿って腋窩領域で内側に折れ曲がります。[ 15 ]
ヴァンヌスはヴァンナルヒダによって区切られており、ヴァンナルヒダは通常、後肘と第 1 ヴァンナル脈の間にあります。直翅目では、通常この位置にあります。しかし、カミキリムシ科の前翅では、この部分のヒダは後肘の直前にしかありません。カワセミ目では、ヴァンナルヒダは後肘の後方にありますが、近位では第 1 ヴァンナル脈の基部を横切ります。セミでは、ヴァンナルヒダは第 1 ヴァンナル脈 (lV) のすぐ後ろにあります。ただし、ヴァンナルヒダの実際の位置のこれらの小さな変動は、翅の屈曲において、屈筋小板 (3Ax) によって制御されるヴァンナル脈の動作の統一性には影響しません。ほとんどの直翅目の後翅では、二次分枝静脈がヴァンナルヒダに肋骨を形成します。ヴァンヌスは通常三角形で、その脈は典型的には第3腋窩から扇の肋のように広がっている。ヴァンヌスの脈の中には枝分かれしているものもあり、二次脈が一次脈と交互に並んでいることもある。ヴァンヌス領域は通常後翅で最も発達しており、プレコプター目や直翅目のように、後翅で拡大して支持面を形成することがある。アカビタイ科の後翅の大きな扇状の拡大は明らかにヴァンヌス領域である。なぜなら、その脈は全て翅基部の第3腋窩小板で支えられているからである。ただし、Martynov (1925) はアカビタイ科の扇状領域のほとんどを翅の頬部領域としている。アカビタイ科の翅の真の頬部は、最後のヴァンヌス脈の小さな膜状 (Ju) 中節によってのみ表される。他の多翅目昆虫、例えばカマキリ科では、羽趾節がより発達している。羽幅の狭い高等昆虫の多くでは、羽趾節が縮小し、羽趾節は消失するが、そのような場合でも、屈曲した羽は後肘部と第一羽趾節の間の線に沿って曲がることがある。[ 15 ]
頬部(ネアラ)は、通常、羽の基部に近い小さな膜状の領域で、いくつかの小さな不規則な静脈状の肥厚によって強化されています。しかし、よく発達すると、翼の独立した部分となり、1つまたは2つの頬静脈を含むこともあります。前翅の頬部が自由葉として発達すると、後翅の上腕角の下に突出し、両翅を繋ぐ役割を果たします。鱗翅目イチジク科では、頬部は長い指状の葉を持ちます。頬部は、明らかに翼の二次的で最近発達した部分であるため、ネアラ(「新しい翼」)と呼ばれています。[ 15 ]
腋窩部は腋窩小板を含む領域で、一般的に不等辺三角形の形をしている。三角形の底辺 (ab) は翼と体のヒンジであり、頂点 (c) は第3腋窩小板の遠位端であり、長い辺は頂点より前方にある。三角形の前側の点 d は、橈骨静脈と第2腋窩小板の接合点を示す。点 d と点 c の間の線は基底襞 (bf)、すなわち中肘部基底部の翼の襞である。[ 15 ]末端は翼の外側の縁であり、頂点と後角または臀角の間にある。[ 17 ] [ 18 ]
一部の双翅目昆虫の翼基部の後方角には、膜状の葉(鱗片またはカリプテルス)が 1 対あり、これらは小羽(alula)と呼ばれています。イエバエでは小羽はよく発達しています。外側の鱗片(c)は、第 3 腋窩小板(3Ax)の後ろの翼基部から発生し、明らかに他の昆虫の頬葉(A、D)を反映しています。より大きな内側の鱗片(d)は、翼節の背板の後盤縁から発生し、托卵の上にフードのような保護的な天蓋を形成します。翼を屈曲させた状態では、小羽の外側の鱗片は内側の鱗片の上で上下逆さまになり、内側の鱗片は翼の動きに影響されません。多くの双翅目昆虫では、単一の羽側脈の後ろにある翅膜の肛門領域に深い切れ込みがあり、小翼の外側鱗片より遠位に近位翼葉が形成される。[ 15 ]
関節

羽のさまざまな動き、特に静止時に羽を背中で水平に折り曲げる昆虫では、羽の基部の関節構造が羽と体の単なる蝶番構造よりも複雑であることが求められる。それぞれの羽は膜状の基底部で体に取り付けられているが、関節膜には多数の小さな関節小板があり、総称して翅翼板と呼ばれる。翅翼板には、肋静脈基部の前方上腕骨板、肋下静脈、橈静脈、前静脈に関連する腋窩静脈群 (Ax)、および中肘部基部の2つのあまり明確でない正中板 (m、m') が含まれる。腋窩静脈は、羽を屈曲させる昆虫でのみ特異的に発達しており、そこでは側膜から発生する屈筋によって操作される羽の屈曲機構を構成している。翼基部の特徴として、上腕骨板近位の関節部前縁に小さな葉状部が存在する。一部の昆虫の前翅では、この葉状部は大きく平坦な鱗片状の膜(テグラ)に発達し、翼基部に重なる。関節膜の後部では、しばしば翼と体の間に広い葉状部を形成し、その縁は一般に厚く波状になっており、靭帯のような外観を呈している。この腋窩索は、翼を担う背板の後縁板襞と内側で連続している。[ 15 ]
羽を曲げる昆虫の羽根の基部の関節小板、または翅目と、その体および羽脈との関係を図式で示すと、次のようになります。
- 上腕骨プレート
- 第一腋窩
- 第二腋窩
- 第三腋窩
- 第四腋窩
- 中央板(m、m')
上腕骨板は通常、翼基部の前縁に位置する小さな板状構造で、可動性があり、肋静脈基部と関節を形成している。トンボ目トンボ類の上腕骨板は大きく拡大しており[ 15 ]、上腕骨上板から発生する2つの筋肉が上腕骨板に挿入され、上腕骨上板の縁から発生する2つの筋肉が腋窩板に挿入されている[ 11 ] 。
第一腋窩小板 (lAx) は翼基部の前方蝶番板である。その前部は背板 (ANP) の前胸翼突起に支持され、後部は背板縁と関節する。小板の前端は一般に細い腕として生じ、その頂点 (e) は常に肋下静脈 (Sc) の基部と結合しているが、肋下静脈とは結合していない。小板本体は第二腋窩小板と側方に関節する。第二腋窩小板 (2Ax) は第一腋窩小板よりも形状が多様であるが、その機械的関係は同様に明確である。第一腋窩小板本体の外縁に斜めに蝶番結合し、その前端 (d) には橈側静脈 (R) が常に柔軟に付着している。第二腋窩骨は翼基部の背側と腹側に硬皮化を呈し、その腹面は側板の支点翼突起上に載っている。したがって、第二腋窩骨は翼基部の枢動骨板であり、特に放射状脈を操作している。[ 15 ]
第三腋窩小板 (3Ax) は、翼の関節部後部に位置する。その形状は多種多様で、しばしば不規則であるが、第三腋窩小板は翼屈筋 (D) が付着する小板である。前側では前方 (f) で第二腋窩小板の後端に、後方 (b) で背板後翼突起 (PNP) に、または後者が存在する場合には小さな第四腋窩小板と関節する。遠位側では、第三腋窩小板は、翼の肛門部にある静脈群の基部と常に関連する突起に沿って延長し、ここでは前静脈 (V) と呼ぶ。したがって、第三腋窩小板は通常、翼基部の後方ヒンジプレートであり、前静脈を直接操作する屈筋機構の能動小板である。屈筋(D)の収縮により、第3腋窩骨は前額関節(b, f)を中心に回転し、遠位腕が挙上する。この動きが翼の屈曲を引き起こす。第4腋窩骨板は翼基部の不変の要素ではない。存在する場合、通常は第3腋窩骨板と後胸翼突起の間に介在する小さな板状構造であり、おそらく後者から分離した部分であると考えられる。[ 15 ]
正中板(m、m')もまた、3つの主腋窩板ほど明確に分化していない板状の小板であるが、それでも屈筋装置の重要な要素である。これらは、第2および第3腋窩板の遠位側の翼基部の正中領域に位置し、翼の屈曲時に顕著な凸状の襞を形成する斜線(bf)によって互いに隔てられている。近位板(m)は通常、第3腋窩板の遠位腕に付着しており、おそらく後者の一部とみなされるべきである。遠位板(m')は、明確な小板として常に存在するわけではなく、翼の肘部内側基部の基部全体の硬化として表されることもある。この領域の静脈が基部で明確に区別される場合、それらは外側の正中板と関連している。[ 15 ]
筋肉

昆虫の飛翔を制御する筋肉は、体重の10%から30%を占めることがあります。飛翔を制御する筋肉は、昆虫に見られる2種類の飛翔、すなわち間接飛翔と直接飛翔の形態によって異なります。間接飛翔を行う昆虫は、その名の通り、筋肉が羽ではなく背板に付着しています。筋肉が収縮すると胸郭が歪み、エネルギーが羽に伝達されます。筋肉には2つの「束」があり、背板に平行に伸びる背縦筋と、背板に付着して胸骨まで伸びる背腹筋です。[ 19 ]直接飛翔を行う昆虫では、胸壁である側板から羽の基部にある個々の小板に直接接続されています。翼下筋と基底筋は、翼下筋と基底筋の小板に靭帯で付着しています。ここで、非常に弾力性のある物質であるレジリンが、飛翔筋を翼器官に結びつける靭帯を形成します。
双翅目(ハエ)や膜翅目(スズメバチ)といったより派生した昆虫目では、間接筋が翼胸郭の大部分を占め、羽ばたきの動力源として機能している。背縦筋の収縮は、背背板を大きく反らせ、翼を押し下げる。一方、背腹筋の収縮は、背背板を反対方向に動かす。現存する最も原始的な飛翔昆虫であるカゲロウ類(カゲロウ)とトンボ類(トンボ)は、上下運動に必要な力を生み出す直接筋を用いている。[ 19 ] [ 20 ]
昆虫の翅筋は完全に好気性の組織である。単位タンパク質当たりの燃料と酸素の消費速度は、非常に高濃度で高度に組織化された組織で起こるため、単位体積当たりの定常状態における消費速度は生物学における絶対的な記録となっている。燃料と酸素を豊富に含んだ血液は、大量の拡散によって筋肉に運ばれ、飛行中に消費される高レベルのエネルギーを維持する。多くの翅筋は大型で、長さ10mm、幅2mmにも達する。さらに、双翅目昆虫の中には、繊維が巨大なものもいる。例えば、非常に活動的なルチリア(Rutilia)では、断面積は長さ1800μm、幅500μmを超える。したがって、周囲から消費部位への燃料と酸素の輸送、そして二酸化炭素の逆輸送は、液相輸送と気管系といった複雑な気管系における輸送の両方の観点から、生物学者にとって難題となっている。[ 21 ]
センサー
昆虫の羽には、味覚毛[ 22 ]、機械感覚毛[ 23 ] 、鐘形感覚子[ 24 ] 、および弦音器官[ 25 ]など、いくつかの種類の感覚ニューロンが存在します。これらのセンサーは、効果的な飛行[ 26 ]と毛づくろい[ 27 ]に必要な外部および内部の固有受容フィードバックを神経系に提供します。
連結、折りたたみ、その他の機能
多くの昆虫種では、前翅と後翅が連結しており、前翅と後翅を1枚の大きな翅に結合させることで、飛行時の空気力学的効率を向上させています。最も一般的な連結機構(膜翅目およびトビケラ目)は、後翅の前縁にある一連の小さなフック(「ハムリ」)で、前翅に引っ掛かり、前翅と後翅を連結した状態(有鉤状連結)です。他の昆虫種(例えば、カメムシ目、チョウ目、一部のトビケラ目)では、前翅の頬葉が後翅の一部を覆う(頬葉連結)、前翅と後翅の縁が広く重なる(アンプレクシフォーム連結)、または後翅の剛毛(小帯)が前翅の保持構造または支帯の下に引っかかる(鉤状連結)ことがあります。[ 14 ]:43
ほとんどの昆虫は、静止しているとき、羽を背中に抱えている。これには、翅膜の縦方向の折り畳みと、ときには横方向の折り畳みが伴う。折り畳みは、屈曲線に沿って起こることもある。カブトムシやハサミムシの後翅のように、折り畳み線が横方向のこともあるが、通常は翅の基部に対して放射状であり、隣接する翅の部分が互いの上または下に折り畳まれることを可能にしている。最も一般的な折り畳み線は、第 3 臀脈のすぐ後ろにある頬襞であるが、[ 12 ]ほとんどの新翅目昆虫は、前翅の第 3A 脈のすぐ後ろに頬襞を持っている。これは、後翅にも存在することがある。直翅目やカブトムシのように、後翅の肛門領域が大きい場合、この部分全体が、鎖骨溝の少し後ろにある前襞に沿って翅の前部の下に折り畳まれることがある。さらに、直翅目とカメムシ目では、肛門部は静脈に沿って扇状に折り畳まれており、肛門静脈は襞の頂点で凸状、副静脈は凹状となっている。鎖骨溝と頬骨襞は異なる種間で相同性を持つと考えられるが、前肢襞は分類群によって位置が異なる。この襞は、側板から発生し第3腋窩小板に挿入される筋肉によって生じ、この筋肉が収縮すると、小板は後胸突起および第2腋窩小板との接合点を中心に回転する。[ 11 ]
その結果、第3腋窩小板の遠位腕は上方および内方に回転し、最終的にその位置は完全に逆転する。肛門静脈はこの小板と連結されており、小板が動くと肛門静脈も一緒に運ばれ、昆虫の背中で屈曲する。飛行中のこの筋肉の活動は羽の出力に影響を与えるため、飛行制御においても重要である。直翅目昆虫では、クチクラの弾力性により、羽の羽側部分が静脈に沿って折り畳まれる。その結果、羽を飛行姿勢に動かす際に、この領域を展開するためにエネルギーが消費される。一般的に、羽の伸展は基底小板、あるいは一部の昆虫では下翼小板に付着する筋肉の収縮によって生じると考えられる。[ 11 ]
フライト

飛行機構
比較的大型の昆虫であるカゲロウ類(カゲロウ)とトンボ類(トンボとイトトンボ)の2つのグループは、飛翔筋が羽に直接付着しており、羽の羽ばたき速度は、神経が飛翔筋に命令を送るインパルスの速度よりも速くはない。[ 28 ]他のすべての有翅昆虫は、異なるメカニズムを用いて飛翔する。それは、胸部を振動させる間接的な飛翔筋である。羽の羽ばたき速度は、飛翔筋が神経インパルスを受信する速度よりも速い。このメカニズムは一度進化し、亜綱新翅目( Neoptera )の特徴(共形質)となっている。[ 28 ]
空気力学
昆虫の飛行には、2つの基本的な空気力学的モデルがあります。ほとんどの昆虫は、螺旋状の前縁渦を作り出す方法を用います。[ 29 ] [ 30 ]非常に小さな昆虫の中には、羽根が昆虫の体上でぶつかり合い、その後羽根が離れる「フリング・アンド・クラップ」、つまりワイス・フォッホ機構を用いるものもあります。羽根が開くと空気が吸い込まれ、それぞれの羽根の上に渦が形成されます。この束縛された渦は羽根を横切って移動し、羽根が離れる際に、もう一方の羽根の起点となる渦として作用します。循環と揚力は増加しますが、羽根の摩耗が増します。[ 29 ] [ 30 ]
多くの昆虫は羽を素早く羽ばたかせることでホバリングすることができ、これには揚力だけでなく横方向の安定も必要となる。[ 31 ]
いくつかの昆虫は推進力を使わずに 滑空飛行をします。
進化
約3億5000万年前の石炭紀、まだ二つの主要な陸地しか存在しなかった時代に、昆虫は飛び始めました。しかし、昆虫の羽がどのように、そしてなぜ発達したのかは、石炭紀前期における羽の発達期に関する適切な化石がほとんど存在しないため、十分に解明されていません。昆虫の飛翔の起源については、主に3つの説があります。羽は胸部背板の延長である傍胸葉から発達したという説、カゲロウなどの水生幼虫に見られる可動式の腹部鰓の変形であるという説、そして放熱器として利用されていた胸部突起から発達したという説です。[ 32 ]
化石

デボン紀(4億年前)の化石はすべて羽がないが、石炭紀(3億2000万年前)までに、10を超える属の昆虫が完全に機能する羽を持っていた。2つの時代間の過渡的形態はほとんど保存されていない。最も古い有翅昆虫はこの時代(有翅亜綱)のもので、バッタ目、カゲロウ目、原始的な基幹グループのカゲロウ類、直翅目、およびパレオディクティオプテロイデス科などが含まれる。非常に初期のバッタ目(石炭紀)は、非常に大きな円盤状の前胸板と、はっきりとしたCuP脈(枝分かれしていない羽脈で、鎖骨襞付近にあり、羽の後縁まで達している)を持つ皮質の前翅を持っていた。[ 33 ] : 399 昆虫の化石として最も古いものはデボン紀のRhyniognatha hirstiで、推定3億9600万~4億700万年前のものであるが、この動物は有翼昆虫に特徴的な二顆状の下顎を持っていた。[ 34 ]しかし後に多足動物であった可能性があると考えられるようになった。[ 35 ]
ペルム紀には、トンボ類(トンボ目)が優勢な空中捕食者で、おそらくは陸生昆虫の捕食も支配していた。真のトンボ目はペルム紀に出現し[ 36 ] [ 37 ]、すべて水陸両生である。その原型は最古の有翼化石であり[ 38 ] 、デボン紀まで遡り、他の翼とはあらゆる点で異なっている。[ 39 ]その原型は石炭紀後期にはすでに多くの現代の特徴の始まりを示していた可能性があり、種によっては翼幅が71cmもあったことから、小型脊椎動物を捕食していた可能性もある。[ 37 ]ペルム紀の最も初期の甲虫のような種は、細胞とピットのある尖った革のような前羽を持っていた。半翅目カメムシ類は、前翅に異常な脈を持つアクチノスキチナ属とパラクナイティア属の形で出現し、おそらくブラットプテラ属から分岐したと考えられる。[ 33 ] : 186
オーストラリア(クロスビー山)では、三畳紀の双翅目の一種の大きな一枚の翅(通常の2~6mmではなく10mm)が発見された。このTilliardipteridae科は、多数の「ティプーロイド」の特徴を持つにもかかわらず、凸状の遠位1A到達翅縁の喪失と肛門ループの形成のため、ヘニングセンシュアルのPsychodomorphaに含まれるべきである。[ 40 ]
仮説

- 傍胸部仮説:この仮説は、昆虫の羽が傍胸部葉から発達したと示唆している。傍胸部葉は昆虫の化石に見られる前適応であり、跳躍や落下時の安定性を助けたと考えられている。この仮説を支持するものとして、ほとんどの昆虫が枝登り中に驚くと地面に落下して逃げる傾向があることが挙げられます。このような葉はパラシュートとして機能し、昆虫がより柔らかく着地することを可能にしたと考えられます。この説は、これらの葉が徐々に大きくなり、後期には胸部と関節を形成したと示唆しています。さらに後期には、これらの粗い羽を動かす筋肉が出現します。このモデルは、パラシュートから始まり、滑空、そして最終的に能動飛行へと、羽の有効性が徐々に高まっていくことを示唆しています。しかしながら、羽の関節と筋肉の発達を示す確かな化石証拠が不足していること、そして一見自発的に発達したように見える関節と脈脈の発達が、この説にとって大きな障害となっています。[ 32 ]
- 上鰓鰓仮説:1870年にカール・ゲーゲンバウアーによって初めて提唱されたこの説は、昆虫の羽の起源として、多くの水生昆虫、例えばカゲロウ類に見られる可動式の腹部鰓が考えられた。 [ 41 ]この説によれば、気管鰓は呼吸器系の出口として発達し、時を経て移動手段として変化し、最終的に羽へと進化した。気管鰓には小さな翼片があり、それらは絶えず振動し、独自の小さな直筋を持つ。[ 32 ]
- エンダイト・エグジット仮説:この仮説は、原始的な節足動物の肢の内側と外側にそれぞれ備わっている付属肢であるエンダイトとエグジットの適応に由来する。この仮説は、トゥルーマン[ 42 ]が、1945年にゴールドシュミットが行ったキイロショウジョウバエの研究に基づいて提唱した。この研究では、莢の変異体において、通常の羽が、いくつかの付属肢を備えた三関節脚と解釈されるものの、通常は羽の肋骨面があるはずの足根を欠く形態へと変化する突然変異が見られた。この突然変異は、脚ではなく、背側のエグジットとエンダイトの融合を強く示唆する証拠として再解釈され、付属肢はこの仮説に非常によく合致した。羽の進化に必要な神経支配、関節構造、筋肉組織は、既に莢節に存在している。[ 32 ]
- パラノタプラス脚遺伝子リクルートメント仮説(二重起源仮説としても知られる) :カゲロウ類の化石幼虫は、昆虫の羽の進化的起源という論争の的となっている問題に新たな重要な手がかりを提供した。幼虫の化石発見以前は、パラノタ仮説と脚出口仮説は互いに矛盾する代替説明と考えられてきたが、どちらも化石記録、比較形態学、発生生物学、遺伝学からの一連の証拠によって裏付けられてきた。昆虫の羽の個体発生における脚遺伝子の発現は、昆虫の羽が可動脚付属器(出口)に由来すると提唱する脚出口仮説を支持する決定的な証拠だと広く考えられてきた。しかし、カゲロウ類の幼虫は、カゲロウとその祖先の腹部の鰓(一般に昆虫の羽に対応する構造と考えられている)が、背側の背板内で関節形成していたことを示している。これは現代のカゲロウの幼虫には見られない。なぜなら、それらの腹部の背板と胸板は輪に融合しており、胚発生においても痕跡を残さないからである。幼虫の鰓と翅が対応する(「連続相同」)構造であり、したがって同じ進化的起源を共有している場合、Coxoplectopteraの新しい結果は、古典的な傍胸骨仮説が提唱するように、翅も背板起源であることを証明している。したがって、Staniczek、Bechly、Godunko(2011)[ 32 ] [ 43 ]は、古生物学と発生遺伝学からの一見矛盾する証拠を調和させることができる新しい仮説を提唱した。すなわち、翅は最初、背板(傍胸骨)の硬い突起として発生し、進化の過程で初めて、脚の遺伝子の二次的なリクルートメントを通じて可動性の関節付き付属器となったのである。[ 32 ]より最近の古生代幼虫の羽根の化石分析は、パラノタの要素と節足動物の脚の遺伝子の融合をさらに裏付けています。[ 44 ]
一部のカワゲラに見られるように、羽は当初水面を滑空するために進化したのではないかという説もある。[ 45 ]別の説では、羽は有翅昆虫の姉妹種である無翅類に見られる飛行前の現象である、指向性滑空降下から派生したものであるという。 [ 46 ]最古の飛行生物はトンボに似ており、2対の羽と直接飛行筋を持ち、腹部に羽を折りたたむ能力はなかった。これらの最初の飛行生物から進化した今日のほとんどの昆虫は、1対の羽か、1対として機能し間接飛行筋システムを使用する2対の羽に単純化されている。[ 32 ]
自然淘汰は、羽、制御および感覚システム、および空気力学や運動学に影響するその他すべてのものの改良に非常に大きな役割を果たしてきました。 注目すべき特徴の 1 つは、羽のねじれです。ほとんどの昆虫の羽は、ヘリコプターの羽根のようにねじれており、根元で迎え角が大きくなっています。ねじれは通常 10 ~ 20 度です。このねじれに加えて、羽の表面は必ずしも平らで特徴がないわけではありません。ほとんどの大型昆虫は、羽の断面が翼型に近づくように、脈の間で翼膜が歪んで角度が付いています。したがって、羽の基本的な形状は、迎え角がゼロのときにすでに少量の揚力を発生させることができます。ほとんどの昆虫は、胸部の小さな筋肉(下図) で羽の傾き、硬さ、羽ばたき頻度を調整することにより、羽を制御します。昆虫の中には、飛行には有利ではないが、交尾や防御など他の役割を果たす他の羽の特徴を進化させたものもあります。[ 32 ]
| 翼を休ませる体の進化方法 | ||
| 羽は折り畳まれない(現生始翅目) | 横に広がる(大きな泡) | |
| 背中同士がぶつかり合う(イトトンボ、カゲロウ) | ||
| 折り畳み式(新翅目) | ||
| 羽は折り畳めない(例:カワゲラ) | ||
| 折りたたみ | 扇形(例:スズメバチの前羽) | |
| クロスフォールド(カブトムシの後ろ翼など) | ||
| 折り畳み対象(ハサミムシの後羽など) | ||
特定の生物学的ニッチを占める昆虫の中には、信じられないほどの機動性を必要とする種もいる。狭い空間で餌を見つけ、大型の捕食者から逃げる能力も必要となる。あるいは、昆虫自身が捕食者となり、獲物を捕獲する必要がある場合もある。空気力学的観点から見ると、昆虫の機動性は高い揚力と推力によってもたらされる。典型的な昆虫の飛行体は、最大で体重の3倍の揚力と、最大で体重の5倍の水平推力を得ることができる。昆虫の飛行機構には大きく異なる2つの種類があり、それぞれに長所と短所がある。トンボ類の飛行機構がより原始的だからといって、飛行能力が劣るわけではない。トンボ類は、ある意味では、それ以降に進化したどの昆虫よりも機敏である。[ 32 ]
形態形成
昆虫における翅の発達は、完全変態する内翅亜綱の昆虫においては明確に定義されています。これらの種では、翅は昆虫のライフサイクルにおける蛹期に発達します。しかし、不完全変態する昆虫には蛹期がないため、異なる翅の形態形成過程を辿ると考えられます。半変態昆虫などの昆虫では、翅は外骨格の下にある蕾から始まり、幼虫の最終齢まで露出しません。[ 47 ]
羽芽の最初の兆候は皮下組織の肥厚で、昆虫種では初期の胚やライフサイクルの最も初期の段階で観察できます。胚での形態学的特徴の発達、つまり胚発生の過程で、細胞クラスターが外胚葉の下で成長します。これは発達の後半で、側方外胚葉が背側に成長して風成虫原基を形成した後で発生します。幼虫における羽芽の発達の例は、モンシロチョウ ( Pieris ) の幼虫に見ることができます。第 2 齢では組織芽細胞がより顕著になり、ポケットのような構造を形成します。第 3 齢および第 4 齢では、組織芽細胞はさらに細長くなります。この大きく延長して膨出または突出した部分が羽になります。最終齢(5齢)の終わりまでに、翅は翅ポケットから押し出されますが、前蛹期の間は古い幼虫のクチクラの下に隠れ続けます。蝶が蛹期に入って初めて翅芽が露出し、羽化後まもなく翅は展開し始め、最終的な形状を形成します。[ 47 ]
翅の気管の発達は翅の組織芽細胞が形成される前に始まる。注目すべきは、気管が大きな気管の近くで発達することである。第4齢期には、この気管上皮細胞が大きく肥大して翅芽の空洞にまで伸び、各細胞は密に巻かれた気管を発達させる。各気管は単細胞起源で、最初は細胞内に位置するが、気管は多細胞起源で、各気管の内腔は細胞間に位置する。気管上皮の単一細胞内にそれぞれ巻かれた気管が発達し、続いて気管と気管内腔との連絡が開き、気管が解けて伸び、翅のあらゆる部分に到達する。[ 47 ]
翼芽は、その発達の初期段階では、気管のような特別な呼吸器官を備えていない。この点では、翼芽がまだ一部である皮下組織の他の部分と類似しているからである。組織芽細胞は大きな気管の近くで発達し、その断面は図に示されている。これは、それぞれ第1齢、第2齢、第3齢、第4齢におけるこれらの部分の断面を表している。同時に、気管は解け、翼芽の形成中の脈腔内で束状に伸びる。蛹期の始まりを示す脱皮において、気管は機能を果たすようになる。同時に、幼虫の気管は退化し、その機能は翼気管に置き換えられる。[ 47 ]
命名法
昆虫目の命名法のほとんどは、古代ギリシャ語で翼を意味するπτερόν ( pteron ) に由来し、接尾辞-ptera が付いています。
| 学名 | 言語的語源 | 学名の翻訳 | 英語名 |
|---|---|---|---|
| 不翅目 | ἀν- ( an- )、ではありません。 ἰσο- ( iso- )、等しい、類似した | 不等翼 | トンボ |
| アプテラ | ἀ- ( a- )、ではない | 翼のない | アプテリゴタン族(現在は廃れています) |
| 無翅亜綱 | πτερύγιον (翼状片小翼) ἀ- ( a- )、ではありません | 翼のない | アプテリゴタン族 |
| 甲虫目 | Κολεός (コレオス、シース) | 強化された翼 | カブトムシ |
| 皮膚ダニ目 | Δέρμα (真皮、皮膚、革) | 革の翼 | ハサミムシ |
| ディアファノプテロデア | Διαφανής (ジアファネス、透明または半透明) | 透明な羽根を持つ | ディアファノプテロイデス類 |
| 網翅目 | Δίκτυον ( diktyon、ネットワーク) | 網状の静脈を持つ翼 | ゴキブリ、カマキリ、シロアリ |
| 双翅目 | Δύο-(dyo-、2) | 二つの翼 | ハエ |
| エンビオプテラ | ἐν- ( en、内部; βίος略歴、人生) | 室内に生息する羽のある昆虫 | ウェブスピナー |
| 内翅亜綱 | ἐντός ( entos、内側; πτερύγιον、小さな翼) | 翼の内側 | 完全変態昆虫 |
| カゲロウ目 | ἐφήμερος (約1日分の 月) | 短命の羽虫 | カゲロウ |
| 外鰭綱 | ἔξω ( exo、外部) | 外翼 | 不完全変態を起こす昆虫(幼虫の頃には外部から羽芽が見える) |
| 半翅目 | ἡμι- ( hemi-、半分) | 半羽の昆虫 | 半翅目(カメムシ、ヨコバイ、アブラムシなど) |
| 異翅目 | ἑτερο- (ヘテロ-、異なる) | 異なる翼を持つ | 真のバグ |
| 同翅目 | ὅμο- ( homo-、類似) | 同じ翼を持つ | 現在は廃止 |
| 膜翅目 | ὑμένιον (処女膜、膜) | 薄い膜の羽を持つ昆虫 | ミツバチ、スズメバチ、アリなど。 |
| 等翅目 | ἶσον ( ison、等しい) | 同じ翼を持つ | シロアリ |
| 鱗翅目 | Λεπίς ( lepis、鱗) | 鱗状の翼 | 蝶と蛾 |
| ロンコプター科 | Λόγχη ( lonche , lance) | ランスウィングス | ランスが飛ぶ |
| メコプター | μῆκος (メコス、長さ) | 長い翼 | ハエトリグモなど |
| メガロプテラ | Μεγαλο- (メガロ-、大きい) | 大きな翼 | ドブソンフライ、フィッシュフライ |
| 神経翅目 | νεῦρον (ニューロン、静脈) | 縞模様の翼 | クサカゲロウ、フクロウバエ、アリジゴクなど。 |
| 新翅目 | νέος ( neos、新しい、若い ) | 新しい翼 | カゲロウとトンボを除く、現在生きているすべての飛翔昆虫目を含む |
| 漸新翅目 | ὀλίγον- ( oligon-、少数) νέος ( neosまたは new) | 小さな静脈のある新しい | 新翅目 |
| 直翅目 | ὀρθο (正, まっすぐ ) | 直線翼 | バッタ、キリギリス、コオロギ |
| 古翅目 | Παλαιός ( Palaios-、古い) δίκτυον ( diktyonネットワークの意味) | 古い静脈のある翼 | 原始的な古生代古翅目昆虫 |
| 古翅目 | Παλαιός (パライオス、古い) | 古い翼 | カゲロウ、トンボ、そしていくつかの化石目 |
| 傍翅目 | Παρα- ( Para- ) νέος ( neos、 new ) | 刺すような口器を持つものがほとんどである新翅目の一部 | カメムシ、シラミ、キジラミ、アザミウマ |
| フティラプテラ | Φθείρ ( phtheir、シラミ) ἀ、a-、not | 羽のないシラミ | 動物のシラミ |
| ウミカメ目 | Πλέκειν (プレカイン、フォールド) | 折りたたまれた翼 | カワゲラ |
| 多翅目 | Πολύς (ポリ、多くのνέοςネオが新しい) | たくさんの静脈のある翼 | 半変態的に発達した新翅目 |
| チャタテムシ目 | Ψώχω (プソコ、こする) | こすり合う羽 | キジラミ、チャタテムシ |
| 翼口亜門 | Πτερύγιον (翼状片、翼) | 羽のある昆虫 | 翅亜綱とは異なり、有翅および無翅二次システムを含む |
| ラフィディオプテラ | ῥαφίς ( rhaphis、針) | 針の羽 | ヘビバエ |
| ノミ目 | Σίφων (サイフォン、チューブ) ἀ- またはなし | 翼のないサイフォン | ノミ |
| ストレプシプテラ | Στρέψις (連鎖球菌、向きを変える) | 回転またはねじれた翼 | ねじれた翼を持つ寄生虫 |
| アザミウマ目 | Θύσανοι (ティサノイ、フリンジ) | フリンジウィング | アザミウマ |
| トビケラ類 | Τρίχωμα (トリコーマ、毛) | 毛のある翼 | トビケラ |
| ゾラプテラ | Ζωρός ( zōrosは強いという意味) | 強い翼 | ゾラプテラ類 |
| 接合翅目 | ζεῦγος (ゼウゴスはペアを意味します) | 一対の翼 | イトトンボ |
適応
変化
昆虫の羽は種の識別と分類において基本的なものであり、昆虫を研究する上でこれより重要な構造は他にない。それぞれの目と科の昆虫は、独特の羽の形と特徴を持っている。多くの場合、色や模様の違いによって種同士を区別することができる。例えば、位置だけでも種を識別することはできるが、その程度ははるかに低い。ほとんどの昆虫は休んでいるときに羽を折りたたむが、トンボや一部のイトトンボは羽を水平に広げて休む。一方、トビケラ、カワゲラ、ハンノキバエ、クサカゲロウなどのグループは、背中の上に傾斜した屋根のような羽を持っている。少数の蛾は羽を体に巻き付けるが、多くのハエとほとんどの蝶は背中の上にまっすぐ上向きに羽を閉じる。[ 48 ]
多くの場合、羽の形状は昆虫の飛翔の種類と相関関係にあります。最も飛行能力の高い昆虫は、細長い羽を持つ傾向があります。スズメガ科(スズメガ)の多くの種では、前羽は大きく鋭く尖っており、小さな後羽と共に三角形を形成し、現代の高速飛行機の翼を連想させます。もう一つの、おそらくより重要な相関関係は、筋肉の大きさと力が飛行速度とパワーに関係していることです。力強く飛ぶ昆虫の羽は、飛行時の応力と空気力学に最も適応しています。羽脈は前縁(または「前縁」)に向かって厚く、強く、密集しており、後縁(または「後縁」)に向かって薄く、かつ柔軟です。これにより、昆虫の羽は優れた構造の翼型となり、抗力を最小限に抑えながら推進力と揚力の両方を発揮することができます。[ 48 ]
羽ばたきの頻度は、種間だけでなく個体間でも、また時期によって変動することがあります。一般的に、羽ばたきの頻度は、羽の筋肉の力と負荷抵抗の比率に依存します。羽が大きく体の軽い蝶の羽ばたき頻度は1秒間に4~20回ですが、羽が小さく体の重いハエやハチは1秒間に100回以上、蚊は1秒間に988~1046回も羽ばたきます。飛行についても同様です。昆虫の飛行速度を推定することは一般的に困難ですが、ほとんどの昆虫は自然界では制御された実験よりも速く飛行できると考えられます。[ 48 ]
甲虫目
甲虫類(甲虫)では、機能的な羽は後翅のみである。後翅は鞘翅よりも長く、鞘翅の下で縦横に折り畳まれる。羽は基部を軸として前方に回転し、飛行姿勢をとる。この動作により、羽は縦横に展開する。羽の構造にはバネ機構があり、腹部の動きも利用して羽を折り畳んだ状態を維持する。甲虫の羽の脈は、この折り畳み構造によって縮小・変形しており、その特徴は以下の通りである。[ 49 ]



- 肋骨 (C)、後肋下骨 (ScP) – 前翼縁にあり、長さのほとんどで癒合している。
- 前橈骨 (RA) – 翼の中央を超えて 2 つの枝に分かれます。
- 後橈骨(RP) – 基底部の接続が失われます。
- 後中膜(MP) – 枝分かれした、長くて強い静脈。
- 前肘(CuA)
- 肛門静脈(AA、AP) – 肘の後ろにある静脈で、肛門のひだによって区切られています。
ほとんどの甲虫種では、前羽は変形して硬化し、鞘翅を形成し、その下に折り畳まれた繊細な後羽を保護します。[ 33 ]鞘翅は翼胸部につながっています。これは、翼がつながっている部分(ギリシャ語で「翼」を意味するpteron )に由来しています。鞘翅は飛翔には使われませんが、体の後部を覆い、第2の翼(alae)を保護する役割を果たします。後部の飛翔翅を動かすには、鞘翅を上げなければなりません。甲虫の飛翔翅は脈と交差しており、着地後にはしばしばこれらの脈に沿って折り畳まれ、鞘翅の下に収納されます。一部の甲虫は飛翔能力を失っています。これには、一部のオサムシ科甲虫やゾウムシ科ゾウムシなどが含まれますが、砂漠や洞窟に生息する他の科の種も含まれています。これらの種の多くは、2枚の鞘翅が融合し、腹部を覆う堅固な盾を形成しています。一部の科では、飛翔能力と鞘翅の両方が失われており、最もよく知られている例は、ツチボタル科のホタルで、雌は生涯を通じて幼虫の状態で成長します。 [ 13 ] [ 33 ]
鱗翅目
2対の羽はそれぞれ中節と第三節、つまり中胸と後胸に見られる。より新しい属では第2節の羽ははるかに顕著であるが、より原始的な形態では両節の羽が同様の大きさである。羽は屋根板状に配置された鱗で覆われており、色彩の驚くべき多様性を形成している。中胸部は蛾や蝶を空中で推進するためにより強力な筋肉を持つように進化し、その節の羽はより強い脈構造を持つ。[ 33 ] : 560 最大の上科であるヤガ科では、羽が鼓室または聴覚器官として機能するように改変されている[ 50 ]羽の脈の改変には以下が含まれる: [ 49 ]
- コスタ (C) – 蝶々には見つかりません。
- 肋下節(Sc) +半径1(Sc+R1) - 前翅縁では、長さの大部分で癒合しているか非常に近いが、後翅では癒合しており、上腕部でよく発達している。蝶では肋下節が分岐することは決してない。
- 半径(R2-R5) – 翼の中央を超えると、アゲハチョウ科では半径が最大5つに分岐します。前翅では、最後のRはセセリ科を除くすべての蝶で柄があり、分離しています。
- 半径セクター(Rs) – 後翼内。
- 中層(M1-M3) – 基底部分が失われています。
- 前肘 (CuA1-CuA2) – CuP セクションが失われています。
- 肛門静脈 (A、1A+2A、3A) – 1 本の静脈 A、または 2 本の静脈 1A+2A、3A。
- 上腕静脈–シジミチョウ科を除くほとんどの蝶の後翅には上腕静脈があります。後翅の上腕部は前翅と重なり合う部分が大きくなっています。上腕静脈は後翅の重なり合う部分を強化し、両翅の接合を良好にしています。
チョウ目の羽、胸部頭部、腹部は微細な鱗片で覆われており、この特徴からチョウ目(Lepidoptera)の学名は、古代ギリシャ語で「鱗片」を意味する「lepteron」に由来しています。ほとんどの鱗片は板状、つまり刃状で、小柄が付いていますが、毛状や二次性徴として特殊化したものもあります。[ 51 ]鱗片の内腔または表面は複雑な構造をしています。鱗片に含まれる色素、あるいはその三次元構造によって、鱗片は色彩を帯びます。[ 52 ]鱗片には、断熱、体温調節、滑空飛行の補助など、様々な機能があります。その中で最も重要なのは、鮮やかな模様から曖昧な模様まで、多様な模様があることです。これらの模様は、カモフラージュ、擬態、そして交尾相手を探す際に、チョウ目にとって重要な役割を果たします。[ 51 ]
トンボ目
トンボ類(イトトンボ類とトンボ類)はどちらも2対の羽を持ち、大きさと形はほぼ同じで、色は透明です。トンボとイトトンボの羽には、R+Mを1とすると5本の主脈があり、羽脈は基部で融合しており、静止時には羽を体に折り畳むことができません。これには以下のものも含まれます。[ 49 ]


- コスタ (C) – 翼の前縁にあり、強くて縁辺部で、翼の頂点まで伸びています。
- 肋下脈(Sc) - 2番目の縦脈。枝分かれせず、結節でC脈と合流する。
- 橈側中膜静脈(R+M)は、翼上で最も強い縦静脈で、3番目と4番目の静脈です。R1~R4の枝は翼縁に達し、中膜前部(MA)も翼縁に達します。IR2とIR3は、それぞれR2とR3の後ろにある介在静脈です。
- 肘 (Cu) - 第 5 縦静脈、後肘 (CuP) は分岐せず、翼縁に達します。
- 肛門静脈(A1) – 肘の後ろにある分岐していない静脈。
- 第二主脈が翼の前縁と接する部分に結節が形成され、翼端付近に黒色の翼柱頭が見られる。
主脈と横脈が翅の脈模様を形成する。脈模様は種によって異なる。横脈は非常に多いこともあれば、むしろ少ないこともある。脈模様は種の同定に有用である。[ 49 ]ほぼ全ての不翅目(Anisoptera)は、翅を横向きまたはわずかに下向きに広げて落ち着くが、ほとんどの接合翅目(Zygoptera)は、翅を寄せ集め、背面を向かい合わせにして落ち着く。接合翅目の胸部は非常に斜めであるため、このように持つと、翅は腹部の上部にきちんと沿う。蝶やカゲロウのように、翅は真上に持ち上げられているようには見えない。いくつかの接合翅目(Zygoptera)の科では、静止時に翅は水平に保たれ、不翅目(Anisoptera)の 1 つの属(例えば、Cordulephya、Corduliidae)では、翅は典型的なイトトンボの静止姿勢で保持される。成虫は 2 対の等しいまたはほぼ等しい翅を持つ。主な脈の茎は 5 本しかないように見える。第二主脈(肋下脈)が翼の前縁と接する部分には結節が形成される。ほとんどの科では、翼端近くに目立つ翼柱頭がある。トンボ目(トンボ類)との同定は脈に基づいて行うことができる。混同されやすいのは、翼に多くの横脈を持つクサカゲロウ科(神経翅目)の一部である。20世紀初頭までは、トンボ目はクサカゲロウ科の近縁種とみなされることがよくあり、準神経翅目(Paraneuroptera)という通称が与えられていたが、この2つの目の類似性は完全に表面的なものである。不等翅目(Anisoptera)では後翅が前翅よりも幅が広く、両翅とも横脈が円盤状の細胞を三角形と超三角形に分割している。[ 53 ]
直翅目

直翅目(バッタやコオロギなど)の種は、前翅が硬くて不透明な細長い板状で、通常は静止時に後翅と腹部を覆っている。後翅は板状の膜状で扇状に折り畳まれており、以下の脈状構造を有する。[ 49 ]
- コスタ(C) – 前翅と後翅の前縁にあり、分岐していない。
- 肋下脈(Sc) – 枝分かれしていない第2縦脈。
- 橈骨 (R) – 第 3 の縦脈。前翅と後翅の R に分岐します。
- 前中膜(MA) - 第4縦静脈。基底部で後中膜(MP)として分岐します。
- 肘 (Cu) – 前翅と後翅にある第 5 の縦脈で、翅の基部近くで分岐した CuA と分岐していない CuP に分かれます。
- 肛門静脈(A) – 肘の後ろにある静脈で、分岐せず、前翅に 2 本、後翅に多数あります。
モース(1920)によれば、被蓋静脈の別の命名法は前方から後方に向かって以下のように示される:[ 54 ]:222–224
- 肋静脈– 一部の種では存在しない場合がある
- 縦隔静脈– 肋骨よりも長いことが多い
- 上腕静脈 – 枝を出す蓋部の中で最も太い静脈
- 円板状静脈 - 上腕骨の枝で、それ自体が分岐していることが多い
- 正中静脈– ほぼ同じ長さの静脈に分岐している
- 鉤状静脈 – 不定に存在
- 尺骨静脈– 枝分かれしており、最も後方の部分は後尺骨静脈または下正中静脈と呼ばれる
- 肛門静脈– 分岐なし
- 腋窩静脈– 分岐せず、翼を折り畳んだときに最上部に位置する
モースによれば、蓋部は3つの領域に分けられる:上腕静脈の前方の辺縁または肋骨領域、上腕静脈と後尺骨静脈の間の円板状または正中領域、および肛門静脈によって前方に境界が付けられた肛門または背側領域である。[ 54 ]:224
ファスマトデア

- コスタ (C) – 後翅の前縁にあり、分岐せず、前翅には存在しない。
- 肋下脈(Sc) – 枝分かれしていない第2縦脈。
- 橈骨 (R) – 3 番目の縦脈。後翅で R に分岐し、前翅では分岐しない。
- 前中膜(MA) - 第4縦静脈。基底部で後中膜(MP)として分岐します。
- Cubitus (Cu) – 5 番目の縦静脈、分岐していません。
- 肛門静脈 (A) – 肘の後ろにある、分岐しない静脈。前翅に 2 本、後翅に多数あり、1A-7A は 1 つのグループに、残りは別のグループに分かれています。
ナナフシの前翅は硬くて不透明なテグミナ(羽片)で、短く、静止時には後翅の基部のみを覆う。肋部から肘部にかけての後翅は、前翅と同様に硬くて不透明である。肛門部は広く、膜状で扇状に折り畳まれている。ナナフシの翅脈には分岐がないか、ごくわずかである。[ 49 ]
皮膚ダニ目


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
ハサミムシ目(Dermaptera )、バッタ目(Orthoptera)、カマキリ目( Mantodea ) 、ゴキブリ目(Blattodea )などの他の目には、飛行中に羽ばたかない硬い革のような前翅があり、テグメン(複数形tegmina)、エリトロン(複数形elytra)、または偽エリトロンと呼ばれることもあります。[ 13 ]
半翅目
半翅目(カメムシ科)では、前翅は硬化しているが、甲虫ほどではない。例えば、カメムシの前翅の前部は硬化しているが、後部は膜状である。これらはヘメリトロン(複数形はヘメリトラ)と呼ばれる。これらは半翅目( Heteroptera)にのみ見られ、セミなどの同翅目の翅は典型的には全体が膜状である。セミの前翅と後翅は両方とも膜状である。ほとんどの種はガラス状であるが、不透明な種もいる。セミは飛ぶのが得意ではなく、ほとんどが数秒しか飛べない。飛ぶとき、前翅と後翅は後翅肋と前翅縁に沿った溝状の結合部によって互いに引っ掛け合わされる。ほとんどの種は、次の図に示すように基本的な脈を持っている。[ 49 ]

- コスタ (C) - 前翼の先端縁で、節まで伸び、Sc+R の近くにあります。
- 肋下節+橈側節(Sc+R) - 前翅ではScとRが節に融合する。節付近で橈側節(Rs)が発生し、分岐が解ける。
- 橈骨前部(RA)
- 後橈骨(RP)
- メディア (M) – M1 から M4 に分岐します。
- 前肘部 (CuA) – CuA1 と CuA2 に分岐します。
- 後肘部 (CuP) – 分岐がありません。
- 肛門静脈(A) - 肘の後ろの静脈。1Aと2Aは前翅で融合し、CuPと2Aは折り畳まれている。
また、両方の翼の縁に周囲の静脈と周辺膜があることにも注意してください。
双翅目
双翅目(ハエ目)では、機能的な羽は1対のみで、後側の羽は托托(はんとう)に縮小している。托托はハエが方向感覚や動きを感知するのに役立つだけでなく、ジャイロスコープのような働きをしてバランスを保つのにも役立つ。ハエ目では、羽の最後部が托托と呼ばれるやや厚くなった羽片に変化し、托托を覆う。[ 49 ]

- コスタ (C) – 双翅目では見つかりません。
- 肋下脈(Sc) – 枝分かれせずに主翼脈となった。
- 半径 (R) – R1~R5 に分岐します。
- 中膜(M) - M1~M4に分岐します。
- 前肘(CuA) - 分岐せず、双翅目ではCuPが縮小する。一部の種ではCuAと1Aが分離し、一部の種では翼縁に達すると接合し、一部の種では融合する。
- 肛門静脈 (A) - 肛門静脈は 1A と 2A の 2 つだけ存在し、2A は種によっては区別がつきません。
- 円板細胞 (dc) – ほとんどの種で明確に定義されています。
カブトムシ目
ゴキブリ類(Blattodea )には、前翅(テグメンとも呼ばれる)があり、これは多かれ少なかれ硬結している。これは飛翔に用いられるだけでなく、膜状の後翅を保護する役割も果たす。後翅の脈は前翅とほぼ同じだが、肛門葉は静止時にCuPとIAの間に折り畳まれる。肛門葉は通常、扇状に折り畳まれる。[ 49 ]

- コスタ (C) – 翼の前縁。
- 肋下静脈(Sc) - 2番目の縦静脈で、比較的短い。
- 橈骨(R) - 多数の櫛状枝を持つ第3の縦脈。
- 中脈(M) - 第4縦脈、翼縁に達する。
- 前肘 (CuA) - 二分枝を持つ第 5 縦静脈が蓋の大部分を占めます。
- 後肘 (CuP) – 分岐せず、湾曲しており、翼縁に達します。
- 肛門静脈(A) – 肘の後ろにある静脈。
膜翅目
| スズメバチ科(Vespidae) における縦折りの例 | |
 | 前翅の主折り線が、明るい水平線として半分ほど上に見えています。この線の後ろにある翅の部分は下向きに折り返されています。翅の前縁にある細い帯状の部分は、前方に折り畳まれた最初の強い線の前にあります。 |
 | そのため、静止状態では、外側の内層が翼の強固な外縁を形成し、衝撃吸収材として腹部の側面を保護します。後翼は大部分が前翼に覆われています。 |
{kind=link}
{kind=link}
ハチ目昆虫の成虫には、ハチ、スズメバチ、ミツバチ、非働きアリなどがあり、いずれも2対の膜状の羽を持っています。[ 49 ]
- コスタ(C) – 膜翅目では見られません。
- Subcosta (Sc) – 分岐していない。
- 半径 (R) – R1~R5 に分岐します。
- 中葉 (M) – M は分岐しておらず、前羽では M の長さの一部が R と融合しています。
- Cubitus (CuA) – 分岐なし、膜翅目では CuP は存在しません。
- 肛門静脈 (A) - 肛門静脈は 1A と 2A の 2 つだけ存在し、2A は種によっては区別がつきません。
- 翼の連結– 後翼の前縁にあるフックの列が前翼の後縁に噛み合い、飛行中に翼を強力に連結します。
- 翼の折り畳み線- スズメバチ科を含む一部の種では、休んでいるときに前翼が「翼の折り畳み線」に沿って縦方向に折り畳まれています。
- 翼柱頭– 一部の種に存在します。
後翅の前縁には、鉤状の剛毛(ハムリ)が多数あり、前翅に引っ掛かり、羽をしっかりと固定します。小型の種では片側に2~3本しかハムリがありませんが、大型のハチではかなりの数のハムリがあり、羽を非常にしっかりと固定します。膜翅目昆虫の羽は、他の多くの昆虫と比較して、特に小型種では比較的少ない静脈を有しています。[ 13 ]
他の家族
シロアリは飛行能力が比較的低く、風速2km/h以下の場合は容易に風下に流され、適切な場所に着地するとすぐに羽を脱ぎ捨て、そこで交尾し、湿った木材や土の中に巣を作ろうとする。[ 55 ]ほとんどのシロアリの羽には、前羽の前縁基部に沿って3本の太い脈があり、羽根の先端付近の横脈は角度がついており、台形の巣房を形成している。一方、地下に生息するシロアリの羽には、前羽の前縁に沿って2本の主脈があり、羽根の先端に向かう横脈はこれらの脈に垂直であるため、正方形や長方形の巣房を形成している。[ 56 ]
アザミウマ類、フシバエ科、その他の飛翔性微小昆虫は、前翅と後翅に長い毛の房があり、縁毛羽とも呼ばれる。[ 57 ]一方、トビケラ類は毛に覆われた羽を持ち、前翅と後翅には剛毛が生えている。[ 13 ]
ストレプシプテラ科のオスも、後翅ではなく前翅から進化した托托を持つ。これは、前翅と後翅に托托を持つ双翅目とは対照的に、ストレプシプテラ科のオスは後翅のみが飛行に機能することを意味する。また、カイガラムシ科のオスの後翅は托托に退化しているか、あるいは存在しない。[ 58 ]
参照
- 付属器
- コムストック・ニーダムシステム
- 翼
- 昆虫にインスパイアされたロボット:RoboBee、DelFly
注記
- ^クランプトン、G. (1916). 「パラノータル理論による昆虫の系統学的起源と羽の性質」ニューヨーク昆虫学会誌. 24 (1): 1– 39. JSTOR 25003692 .
- ^ a bロス、アンドリュー (2017). 「昆虫の進化:羽の起源」 . Current Biology . 27 (3): R113– R115. doi : 10.1016/j.cub.2016.12.014 . PMID 28171756 – Web of Science経由.
- ^ a b Averof, Michalis, and SM Cohen. (1997). 「昆虫の羽の進化的起源は先祖の鰓から」Nature 385 (6617): 627– 630. Bibcode : 1997Natur.385..627A . doi : 10.1038/385627a0 . PMID 9024659. S2CID 4257270 – Web of Science経由.
{{cite journal}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ Grodnitsky, Dmitry, L. (1999).昆虫の羽の形と機能:生物学的構造の進化. pp. 82– 83.
{{cite book}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ a b cアレクサンダー、デイビッド、E. (2015). 『翼の上:昆虫、翼竜、鳥類、コウモリ、そして動物の飛翔の進化』オックスフォード大学出版局. pp. 74– 101.
{{cite book}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ Haug, Joachim, C. Haug., RJ Garwood. (2016). 「昆虫の羽の進化と発達 ― 古生代幼虫の新たな知見」 . Biological Reviews . 91 (1): 53– 69. doi : 10.1111/brv.12159 . PMID 25400084. S2CID 21031689 .
{{cite journal}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ Almudi, Isabel; Vizueta, Joel; Wyatt, Christopher DR; de Mendoza, Alex; Marlétaz, Ferdinand; Firbas, Panos N.; Feuda, Roberto; Masiero, Giulio; Medina, Patricia; Alcaina-Caro, Ana; Cruz, Fernando (2020). 「カゲロウの水生および空中生活へのゲノム適応と昆虫の羽の起源」 . Nature Communications . 11 ( 1): 2631. Bibcode : 2020NatCo..11.2631A . doi : 10.1038/s41467-020-16284-8 . ISSN 2041-1723 . PMC 7250882. PMID 32457347 .
- ^ブルース、ヘザー、NHパテル (2020). 「甲殻類の脚パターン形成遺伝子のノックアウトは、昆虫の羽と体壁が古代の脚節から進化したことを示唆している」. Nature Ecology & Evolution . 4 (12): 1703– 1712. Bibcode : 2020NatEE...4.1703B . doi : 10.1038/ s41559-020-01349-0 . PMID 33262517. S2CID 227253368 .
{{cite journal}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ Clark-Hatchel, Courtney (2013). 「コクゾウリムシTribolium castaneumにおける痕跡小胞の機能解析による昆虫の翅の起源に関する知見」 .米国科学アカデミー紀要. 110 (42): 16951– 16956. Bibcode : 2013PNAS..11016951C . doi : 10.1073/pnas.1304332110 . PMC 3801059. PMID 24085843 .
- ^ Prokop, Jakub, Pecharová, M., Nel, A., Hörnschemeyer, T., Krzemińska, E., Krzemiński, W., & Engel, M (2017). 「古生代幼虫の翅パッドは昆虫の翅の起源に関する二重モデルを支持する」 . Current Biology . 27 (2): 263– 269. doi : 10.1016/j.cub.2016.11.021 . PMID 28089512 .
{{cite journal}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ a b c d e fチャップマン, RF (1998). 『昆虫:構造と機能』(第4版)ケンブリッジ、ニューヨーク:ケンブリッジ大学出版局. ISBN 0-521-57048-4。
- ^ a bジリオット、セドリック (1995 年 8 月)。昆虫学(第 2 版)。 Springer-Verlag New York, LLC. ISBN 0-306-44967-6。
- ^ a b c d e Meyer, John R. (2007年1月5日). 「外部解剖学:翼」ノースカロライナ州立大学昆虫学部. 2011年7月16日時点のオリジナルよりアーカイブ。 2011年3月21日閲覧。
- ^ a b c dガラン、PJ; PS、クランストン(2004)。昆虫:昆虫学の概要。英国: ブラックウェル出版。 p. 42.ISBN 1-4051-1113-5。
- ^ a b c d e f g h i j k l m n oスノッドグラス, RE (1993年12月).昆虫形態学の原理. コーネル大学出版局. ISBN 0-8014-8125-2。
- ^ Spieth, HT (1932). 「カゲロウ類の翅脈を研究する新しい方法とそこから得られたいくつかの結果」(PDF) .昆虫学ニュース. 2011年9月30日時点のオリジナル(PDF)からアーカイブ。
- ^ 「昆虫学で使用される用語の説明」 www.gutenberg.org . 2024年1月8日閲覧。
- ^ 「TERMENの定義」 www.merriam-webster.com . 2024年1月8日閲覧。
- ^ a b Knospe, Carl R. (1998年秋). 「昆虫の飛行メカニズム:解剖学と運動学」(PDF) . バージニア大学機械・航空宇宙工学科.
- ^ Weis-Fogh, T (1963年7月). 「昆虫の翅筋における拡散:最も活発な組織として知られる」 . J Exp Biol . 41 (2): 229– 256. doi : 10.1242/jeb.41.2.229 . PMID 14187297 .
- ^ Tiegs, OW (1954年2月). 「昆虫の飛翔筋 - 解剖学と組織学; 横紋筋の構造全般に関する考察」 .ロンドン王立協会哲学論文集. シリーズB, 生物科学. 238 (656): 221– 348. Bibcode : 1955RSPTB.238..221T . doi : 10.1098/rstb.1955.0001 . JSTOR 3739600 .
- ^ He, Zhe; Luo, Yichen; Shang, Xueying; Sun, Jennifer S.; Carlson, John R. (2019年5月21日). 「ショウジョウバエの翅の化学感覚器はイオンチャネル型フェロモン受容体候補を発現する」 . PLOS Biology . 17 (5) e2006619. doi : 10.1371/journal.pbio.2006619 . PMC 6528970 .
- ^ Valmalette, JC, Raad, H., Qiu, N., Ohara, S., Capovilla, M. and Robichon, A., 2015. ショウジョウバエの羽における味覚化学感覚毛と気管のナノ構造. Scientific reports, 5(1), pp.1-11.
- ^ Dinges, GF, Chockley, AS, Bockemühl, T., Ito, K., Blanke, A. and Büschges, A., 2021. Drosophila melanogasterにおける鐘形感覚子の位置と配置. Journal of Comparative Neurology, 529(4), pp.905-925.
- ^ Field, LHおよびMatheson, T., 1998. 昆虫の弦音器官. 昆虫生理学の進歩(第27巻、pp. 1-228). アカデミックプレス.
- ^ Wolf, H., 1993. バッタのテグラ:飛行リズム生成、羽運動制御、空気力生成における意義. 実験生物学ジャーナル, 182(1), pp.229-253.
- ^ Zhang, N.とSimpson, JH, 2022.「一対の交連指令ニューロンがショウジョウバエの羽のグルーミングを誘導する」Iscience, 25(2), p.103792.
- ^ a b Chapman, AD (2006).オーストラリアと世界の生物種の数. キャンベラ:オーストラリア生物資源研究. 60ページ. ISBN 978-0-642-56850-2. 2009年5月19日時点のオリジナルよりアーカイブ。2012年6月18日閲覧。
{{cite book}}: CS1 maint: publisher location (link) - ^ a b Wang, Z. Jane (2005). 「昆虫の飛行の解剖」(PDF) . Annual Review of Fluid Mechanics . 37 (1). Annual Reviews: 183– 210. Bibcode : 2005AnRFM..37..183W . doi : 10.1146/annurev.fluid.36.050802.121940 . 2014年1月16日時点のオリジナル(PDF)からアーカイブ。2011年3月30日閲覧。
- ^ a b Sanjay P. Sane (2003). 「昆虫の飛行の空気力学」(PDF) . The Journal of Experimental Biology . 206 (23): 4191– 4208. doi : 10.1242/jeb.00663 . PMID 14581590. S2CID 17453426 .
- ^ Davidovits, Paul (2008). Physics in Biology and Medicine . Academic Press. pp. 78– 79. ISBN 978-0-12-369411-9。
- ^ a b c d e f g h iグリマルディ、デイビッド、エンゲル、マイケル・S. (2005). 『昆虫の進化』ニューヨーク、NY: ケンブリッジ大学出版局.
- ^ a b c d e Powell, Jerry A. (2009). 「Coleoptera」 . Resh, Vincent H.; Cardé, Ring T. (編). Encyclopedia of Insects (2 (illustrated) ed.). Academic Press. p. 1132. ISBN 978-0-12-374144-8。
- ^ Michael S. Engel; David A. Grimaldi (2004). 「最古の昆虫に新たな光」. Nature . 427 ( 6975): 627– 630. Bibcode : 2004Natur.427..627E . doi : 10.1038/nature02291 . PMID 14961119. S2CID 4431205 .
- ^ハウグ, カロリン; ハウグ, ヨアヒム T. (2017-05-30). 「推定最古の飛翔昆虫:多足動物の可能性が高い?」 . PeerJ . 5 e3402 . doi : 10.7717/peerj.3402 . ISSN 2167-8359 . PMC 5452959. PMID 28584727 .
- ^ Grzimek HC Bernhard (1975) Grzimekの動物百科事典第22巻昆虫。Van Nostrand Reinhold Co. NY。
- ^ a b Riek EF Kukalova-Peck J (1984). 「アルゼンチン産上部石炭紀前期の化石に基づくトンボの翅脈の新たな解釈(昆虫綱:トンボ目および翼状部翅の基本的特徴)」Can. J. Zool . 62 (6): 1150– 1160. doi : 10.1139/z84-166 .
- ^ Wakeling JM Ellington CP (1997). 「トンボの飛行IIIにおける揚力と出力要件」 . Journal of Experimental Biology . 200 (Pt 3): 583–600 (589). doi : 10.1242/jeb.200.3.583 . PMID 9318294 .
- ^松田 良治 (1970). 「昆虫胸部の形態と進化」. Mem. Entomol. Soc. Can . 102 (76): 1– 431. doi : 10.4039/entm10276fv .
- ^ VA Blagoderov; ED Lukashevich; MB Mostovski (2002). 「Order Diptera Linné, 1758. The true flies.」 AP Rasnitsyn; DLJ Quicke (eds.). History of Insects . Kluwer Academic Publishers . ISBN 1-4020-0026-X。
- ^カール、ゲーゲンバウア (1870)。Grundzüge der vergleichenden Anatomy。 W.エンゲルマン。
- ^ Trueman JWH (1990)、コメント:昆虫の羽の進化:肢体出口と末端モデル Canadian Journal of Zoology
- ^ Staniczek, AH; Bechly, G. & Godunko, RJ (2011). 「Coxoplectoptera, a new fossil order of Palaeoptera (Arthropoda: Insecta), with comments on the phylogeny of the stem group of mayflies (Ephemeroptera)」(PDF) . Insect Systematics & Evolution . 42 (2): 101– 138. doi : 10.1163/187631211X578406 . S2CID 4986911. 2011年10月3日時点のオリジナル(PDF)からアーカイブ。
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^プロコップ、ヤクブ;ペチャロバ、マルティナ。ネル、アンドレ。ヘルンシェマイヤー、トーマス;クシェミンスカ、エヴァ。クシェミンスキ、ヴィエスワフ。マイケル・S・エンゲル(2017年1月)。「古生代の若虫の羽パッドは昆虫の羽の起源のデュアルモデルをサポートします。 」現在の生物学。27 (2): 263–269。土井: 10.1016/j.cub.2016.11.021。PMID 28089512。
- ^ Adrian LR Thomas; R. Åke Norberg (1996). 「表面をかすめる — 昆虫の飛翔の起源?」Trends in Ecology & Evolution . 11 (5): 187– 188. doi : 10.1016/0169-5347(96)30022-0 . PMID 21237803 .
- ^ Yanoviak SP, Kaspari M, Dudley R (2009). 「滑空する六脚動物と昆虫の空中行動の起源」 . Biology Letters . 5 (4): 510–2 . doi : 10.1098/rsbl.2009.0029 . PMC 2781901. PMID 19324632 .
- ^ a b c d H. Comstock, Henry (1918). The Wings of Insects . Ithaca, NY: The Comstack Publishing Company. p. 114 .
- ^ a b c「昆虫の羽根全般」昆虫の空気力学Cislunar Aerospace. 1997年. 2011年3月28日閲覧。
- ^ a b c d e f g h i j Chew, Peter (2009年5月9日). 「昆虫の羽」 . Brisbane Insects and Spiders . 2011年3月21日閲覧。
- ^ Scoble, MJ. (1992).鱗翅目昆虫:形態、機能、多様性. オックスフォード大学出版局. ISBN 978-1-4020-6242-1。
- ^ a b Scoble (1995). Section Scales , (pp 63 – 66).
- ^ Vukusic, P. (2006). 「鱗翅目昆虫の構造色」(PDF) . Current Biology . 16 (16): R621–3. doi : 10.1016/j.cub.2006.07.040 . PMID 16920604. S2CID 52828850. 2010年11月11日閲覧.
- ^ Trueman, John WH; Richard J. Rowe (2009年10月16日). 「トンボ類. トンボとイトトンボ」 . Tolweb.org . 2011年3月21日閲覧。
- ^ a bモース、アルバート・P. (1920). 「ニューイングランドの直翅目昆虫マニュアル(イナゴ、バッタ、コオロギ、およびその仲間を含む)」(PDF) .ボストン自然史協会紀要. 35 (6): 197– 556. 2025年11月5日閲覧– The Orthopterists' Society経由.
- ^ Abe T., Bignell DE; Hiragana M. (2000).シロアリ:進化、社会性、共生、生態、エコラボ. Kluwer academic publishers. ISBN 0-7923-6361-2。
- ^ 「シロアリ」テキサス・アグリライフ・エクステンション・サービス。 2011年4月13日時点のオリジナルよりアーカイブ。
- ^ Polilov, Alexey A.; Reshetnikova, Natalia I.; Petrov, Pyotr N.; Farisenkov, Sergey E. (2019年1月). 「羽根つき甲虫(甲虫目:羽根つき甲虫科)の翅の形態:小型化と機能的スケーリング解析に関連する特徴」 .節足動物の構造と発達. 特集:汎節足動物の小型化. 48 : 56–70 . Bibcode : 2019ArtSD..48...56P . doi : 10.1016/j.asd.2019.01.003 .
- ^ Camacho, Ernesto Robayo; Chong, Juang-Horng (2015). 「軟質カイガラムシ類(半翅目:コナカイガラムシ科)の一般生物学と最新の管理アプローチ」(PDF) . Journal of Integrated Pest Management . 6 (1): 17. doi : 10.1093 /jipm/pmv016 . ISSN 2155-7470 . PMC 4725186. PMID 26823990 .
参考文献
- トリプルホーン、チャールズ・A.、ジョンソン・ノーマン・F. (2005).ボラーとデロングによる昆虫学入門(第7版). トムソン・ブルックス/コール. ISBN 0-03-096835-6。
外部リンク
