This article needs additional citations for verification. (September 2025) |

{kind=link}
This article may require cleanup to meet Wikipedia's quality standards. The specific problem is: redundant sections need rewriting. (September 2025) |
| ラティメリア | |
|---|---|

| |
| インドネシアシーラカンス(Latimeria menadoensis) | |

| |
| 科学的分類 | |
| 界: | |
| 動物界 | 門: |
| 脊索動物門 | 綱: |
| アクチニスト綱 | 放線虫綱 |
| シーラカンス目 | シーラカンス目 |
| ラティメリオイデ上科 | ラティメリオイデ |
| ラティメリア科 | ラティメリオイデ科 |
| Latimeria Smith , 1939 | タイプ種 |
| 種 | |
| ラティメリア・カルムナエ | |
| ラティメリア・カルムナエ | |
| |

| |
| シノニム[ 1 ] | |
| シノニム[1] | |
| |
{kind=link}
{kind=link}
{kind=link}
ラティメリア属は、シーラカンスの現生種が2種のみ含まれる希少な魚類の属です。現生種は、西インド洋シーラカンス( Latimeria chalumnae)とインドネシアシーラカンス( Latimeria menadoensis)の2種です。これらは、肉鰭綱(肉鰭類および四肢動物)の中で最も古い系統に属しており、一般的な条鰭類や軟骨魚類よりも、肺魚や四肢動物(両生類、爬虫類、哺乳類)に近いと考えられています。
インド洋とインドネシアの海岸沿いに生息しています。[2] [3]シーラカンスは2種しか知られておらず、どちらも絶滅の危機に瀕しているため、世界で最も絶滅の危機に瀕している動物の属の1つです。西インド洋シーラカンスは絶滅危惧種に指定されています。[4]
説明
{kind=link}

ラティメリア属のシーラカンスは、体長2メートル(6.6フィート)以上、体重約90キログラム(200ポンド)にまで成長する、大きくふっくらとした肉鰭類の魚です。[5]鱗の年輪の分析に基づくと、最大100年生きると推定され、55歳前後で成熟に達します。[6]知られている最高齢の標本は、1960年に捕獲された時点で84歳でした。[7]
科学者たちは、シーラカンスの耳の骨(耳石)にある年輪に基づいて、個々のシーラカンスは80年から100年も生きる可能性があると推測しています。シーラカンスは海面下700メートル(2300フィート)の深さに生息しますが、より一般的には90~200メートル(300~660フィート)の深さで見つかります
ラティメリア・カルムナエの現生個体は深い青色をしており、これはおそらく獲物からカモフラージュするためでしょう。一方、インドネシア産の種(L. menadoensis)は茶色です。
解剖学と生理学
シーラカンスの目は非常に敏感で、タペタム(透明体)を持っています。シーラカンスは日中に捕獲されることはほとんどありませんが、月の満ち欠けを問わず捕獲されています。シーラカンスの目には、網膜にある受容体である桿体が多くあり、薄暗い場所でも物を見るのに役立ちます。桿体とタペタムは一緒になって、暗い水中で魚がよりよく見えるようにするのに役立ちます。目は、主に短波長を吸収する桿体によって、暗い場所でも見やすく慣れています。シーラカンスの視覚は、主に青方偏移した色覚能力へと進化しました。[8] 偽上顎襞は口を囲み、上顎骨を置き換えていますが、シーラカンスにはこの構造がありません前上顎骨と側頭吻骨の間には、2つの鼻孔と4つの外部開口部が見られる。鼻嚢は他の多くの魚類のものと類似しており、内鼻孔は存在しない。シーラカンスの吻部器官は脳蓋の篩骨部内にあり、周囲に通じる3つの保護されていない開口部を有し、シーラカンスの側方感覚系の一部として利用されている。[9] : 14–16, 19 シーラカンスの聴覚受容は内耳によって行われ、これは四肢動物のものと非常に類似しており、基底乳頭に分類される。[10]
シーラカンスの心臓は、ほとんどの現生魚類とは異なり、心臓室が直管状に配列している。シーラカンスの脳蓋は98.5%が脂肪で満たされており、脳組織は脳蓋のわずか1.5%にしか含まれていない。シーラカンスの頬部は、鰓蓋骨が非常に小さく、大きな軟部組織からなる鰓蓋弁を備えているという点で独特である。気門室は存在するが、気門は閉じており、発達過程においては決して開かない。[11] [9] : 15 [12]現生シーラカンスに特有のものとして、「脂肪肺」、すなわち脂肪で満たされた単葉の退化した 肺の存在がある。これは他の魚類の浮袋に相同である。浮力制御のための脂肪器官が同時に発達したことは、深海生息域への特異な特殊化を示唆している。成体の標本では、退化した肺の周りには小さくて硬いが柔軟性のある板がありますが、脂肪器官の周りにはありません。これらの板は肺の容積を調節する機能を持っていた可能性が高いです。[13]脂肪器官の大きさから、研究者たちはこれが腎臓の異常な位置の原因であると推測しています。2つの腎臓は1つに融合しており、[14]腹腔内の腹側、総排泄腔の後方に位置しています。[15]
科学的研究によると、シーラカンスは冷たく酸素が豊富な水中に留まらなければ、血液が十分な酸素を吸収できないことが示唆されています。[16]この魚は環境に非常によく適応しているようで、これが既知の脊椎動物の中で最も遅いゲノム進化を持つ理由の1つと考えられています。[17]
生物学
シーラカンスは夜行性の魚食性漂流魚である。[18]シーラカンスは日和見主義的な捕食者であり、コウイカ、イカ類、タシギウオ、小型のサメ、および生息地である深い岩礁や火山斜面に生息する他の魚類を狩る。シーラカンスは獲物を見つけるために頭を下に向けたり、後ろ向きに、または腹を上にして泳ぐことも知られているが、これはおそらく吻側腺を使用しているためである。移動するために、彼らは上昇流または下降流と漂流を利用するのが最も一般的である。彼らの一対の鰭は水中での動きを安定させる。海底にいる間は、彼らはいかなる移動にも一対の鰭を使用しない。シーラカンスは素早いスタートのために尾鰭で推進力を生み出す。鰭が豊富なため、シーラカンスは機動性が高く、水中でほぼどの方向にも体を向けることができる。シーラカンスは逆立ちをしたり、腹ばいで泳いだりする姿が目撃されています。吻側器官はシーラカンスに電気受容を与え、障害物を回避するのに役立つと考えられています。[18]
彼らは「受動的な漂流摂食者」であり、最小限の自力でゆっくりと流れに沿って漂い、遭遇した獲物を食べます。[19] [20]シーラカンスは、暗い場所でも近くの獲物を見つけるために、吻側器官を電気受容にも使用しています。[21]
{kind=link}

シーラカンスは同種の個体と遭遇するとかなり穏やかです。身体接触は避けますが、接触した場合はすぐに退却します。外部の潜在的な捕食者(例えば潜水艇)に近づくとパニック逃走反応を示すため、シーラカンスは大型の深海捕食者の餌食である可能性が高いと考えられます。シーラカンスにはサメの噛み跡が見られ、サメはシーラカンスが生息する地域ではよく見られます。[20] 14種類のシーラカンス酵素の 電気泳動試験では、シーラカンスの個体群間で遺伝的多様性がほとんどないことが示されています。捕獲された魚の中には、オスとメスがほぼ同数いました。[9] : 38–40 個体数の推定値は、個体群あたり210個体から500個体の範囲です。[9] : 39 [22]シーラカンスは個体ごとに色の模様があるため、科学者たちは電気通信によって他のシーラカンスを認識していると考えています。[20]
南アフリカのソドワナ湾付近に生息するシーラカンスは、日中は水深90~150m(300~490フィート)の洞窟で休息しますが、夜間に狩りをするときは分散して55m(180フィート)ほどの浅い深さまで泳ぎます。水深は、非常に薄暗い光と、さらに重要なのは14~22℃(57~72°F)の水温を必要とすることほど重要ではありません。彼らはこれらの条件を見つけるために浮上したり沈んだりします。血液が鰓を通して水から吸収できる酸素の量は、水温に依存します。
科学者たちは、この魚がこれほど成功している理由の一つは、標本が代謝を自由に遅くし、人の少ない深みに沈み、一種の冬眠モードで栄養所要量を最小限に抑えることができるためだと考えています。
繁殖
{kind=link}

{kind=link}

シーラカンスは卵胎生で、メスは受精卵を体内に保持し、5年間の妊娠期間中に胚が成長します。一般的にメスはオスよりも大きく、鱗や総排泄腔周囲の皮膚のひだは異なります。オスのシーラカンスには明確な交尾器官はなく、総排泄腔のみがあり、そこには勃起性カルンクルに囲まれた泌尿生殖乳頭があります。総排泄腔は交尾器官として機能するために反転すると考えられています。[9] : 27 [23]シーラカンスの卵は大きく、薄い膜層で保護されています。胚はメスの体内で孵化し、最終的には生きたまま生まれますが、これは魚類では珍しいことですこの事実は、1975年にアメリカ自然史博物館が最初のシーラカンス標本を解剖し、5つの胚を妊娠しているのを発見した際に初めて明らかになった。[24]若いシーラカンスは成体に似ているが、主な違いは外側に卵黄嚢があること、体の大きさに比べて目が大きいこと、そして体の下向きの傾斜が顕著である。若いシーラカンスの幅広い卵黄嚢は腹鰭の下に垂れ下がっている。幼体の鱗と鰭は完全に成熟しているが、成熟期に獲得する歯鰭は欠いている。[23]
メスのシーラカンスは、「パップ」と呼ばれる幼生を一度に5匹から25匹の群れで出産します。幼生は生後すぐに自力で生存することができます。繁殖行動はよく分かっていませんが、 20歳になるまで性成熟しないと考えられています。妊娠期間は13~15ヶ月と考えられていましたが、2021年に行われた研究では、妊娠期間は最大5年であることが示唆されており、 [25] [26]これは、以前の記録保持者である深海ラブカザメよりも1.5年長いものです。[27]
2頭のシーラカンスの雌の体内の胚の父性を評価し、それぞれの卵は1頭の雄によって生まれたことが示されました。[28]これは、雌が一夫多妻制、つまり1頭の雄とのみ交尾することを意味している可能性があります。一夫多妻制、つまり雌が複数の雄と交尾することは、植物と動物の両方に共通しており、有利な場合もあります(例:不妊または不適合な配偶者との交尾に対する保険)が、コストも伴います(感染リスクの増加、捕食者の餌食になる危険性、新しい雄を探す際のエネルギー投入の増加)。[要出典]
進化的関係
{kind=link}
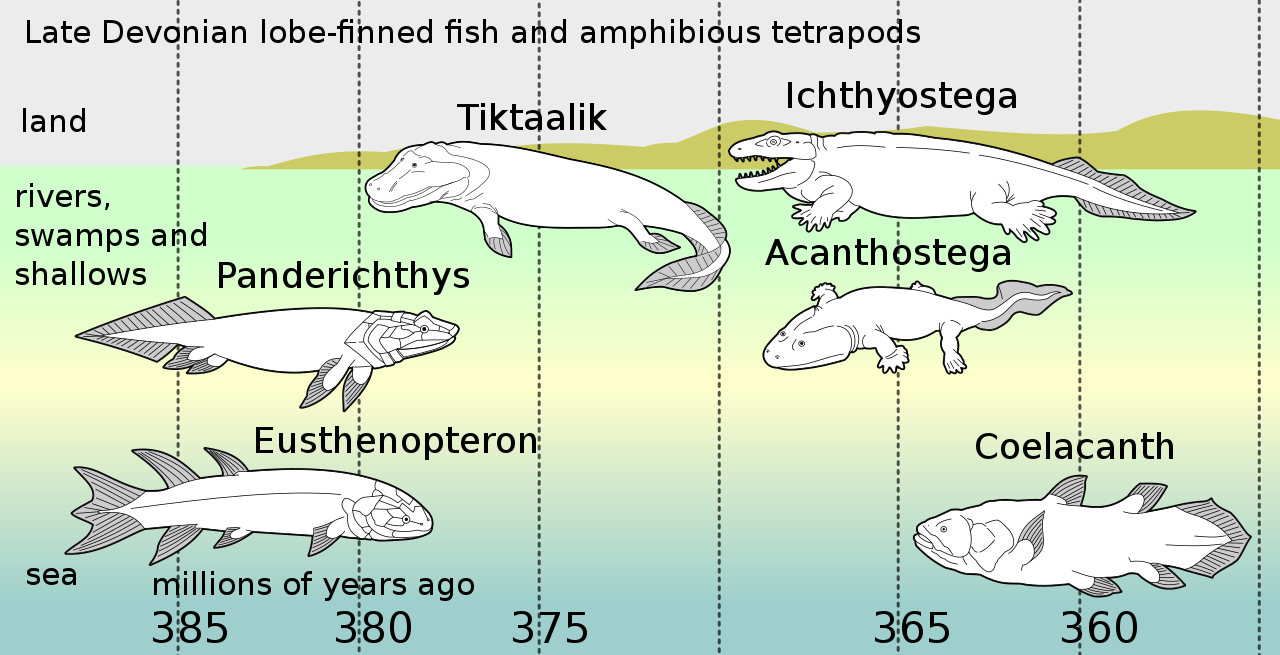
ラティメリア属は、三畳紀前期に初めて出現したシーラカンスの一種であるラティメリア科のタイプ属です。ラティメリア科の中では、分岐論的解析により、ヨーロッパのジュラ紀後期に知られるスウェンジア属に最も近縁であることが判明しており、最も近い近縁種から1億5000万年以上の長いゴースト系統を残しています。[29]
インドネシア東部、西パプア州ワイゲオで回収されたインドネシア産シーラカンスの標本の分析によると、インドネシア産シーラカンスには別の系統が存在する可能性があり、2つの系統は1300万年前に分岐した可能性があることが示唆されています。この新しい系統が亜種か新種かはまだ判明していません。[30]
遺伝学
2013年、ある研究グループが科学誌「ネイチャー」にシーラカンスのゲノム配列を発表しました。[31]
シーラカンスは、葉状の鰭やその他の特徴から、かつては四肢動物以外の肉鰭類の中で最も新しい分岐である可能性があると仮説が立てられていました。 [ 10 ] [32]しかし、シーラカンスの全ゲノム配列を解読した後、肺魚の方が四肢動物に近縁であることが発見されました。シーラカンスとリピディスト類(肺魚と四肢動物の同族)は、肺魚が陸生に移行する前からすでに分岐していました。[33]
ゲノム配列解読から得られたもう一つの重要な発見は、シーラカンスが現在も進化を続けているということです。現生シーラカンスと絶滅シーラカンスの表現型の類似性は、これらの生物が形態的分岐を起こすための進化圧力は限られていることを示唆していますが、測定可能な遺伝的分岐は起こっています。タンパク質コード領域は他の肉鰭綱よりもはるかに低い置換率で進化していることが以前の研究で示されているにもかかわらず(この分類群の現生種と化石種の間で観察される表現型の停滞と一致する)、より高い転移因子活性を受ける非コード領域は、現生の2つのシーラカンス種間でも顕著な分岐を示しています。[31]これは、イプシロンレトロウイルスファミリーのシーラカンス特異的な内在性レトロウイルスによって部分的に促進されています。[34]
分布
シーラカンスの現在の生息域は主にアフリカ東海岸ですが、インドネシア沖でもラティメリア・メナドエンシスが発見されています。シーラカンスはケニア、タンザニア、モザンビーク、南アフリカ、マダガスカル、コモロ、インドネシアの海域で発見されています。[3]これまでに捕獲されたラティメリア・カルムナエの標本のほとんどは、コモロ諸島(インド洋)のグランドコモロ島とアンジュアン島周辺で捕獲されています。他の地域でもラティメリア・カルムナエが捕獲された事例はありますが、アミノ酸配列解析の結果、コモロ島とアンジュアン島周辺で発見されたものと大きな違いは見られませんでした。これらの少数の個体は迷子と考えられるものの、マダガスカル沖でシーラカンスが捕獲されたという報告はいくつかあります。このことから科学者たちは、 Latimeria chalumnaeシーラカンスの固有分布域は、コモロ諸島からアフリカ東海岸に沿ってマダガスカル西海岸を過ぎて南アフリカの海岸線まで広がっていると考えています。[9] : 13, 32, 35 タンザニア南部の海岸で捕獲されたシーラカンスのミトコンドリアDNA配列は、約20万年前に2つの個体群が分岐したことを示唆しています。これは、コモロ個体群が主要個体群であり、他の個体は最近の分岐であるという説に反証となる可能性があります。[35]生きた標本は2019年11月にクワズール・ナタール州南海岸のウムズンベ村の沖合69メートル、イシマンガリソ湿地公園の南325キロで目撃され、ビデオに記録されました。これは最初の発見以来最も南下した記録であり、ディープガット海峡の54メートル(177フィート)に次いで2番目に浅い記録です。これらの目撃情報は、少なくとも生息域の南端では、これまで考えられていたよりも浅い場所に生息している可能性を示唆しています。南端では、より冷たく、より酸素に富んだ水が浅い深度で利用できるからです。[36]
インドネシアシーラカンス(Latimeria menadoensis )の分布域は、インドネシア、スラウェシ島マナド・トゥア島沖、セレベス海であると考えられています。[2]シーラカンスをこれらの地域に限定する主な要因は、餌と温度の制限、そして漂流摂食に適した洞窟や割れ目などの生態学的要件です。[19]潜水艇を使用した研究チームは、スラウェシ海とパプアのビアク島海域でこの魚の生目撃を記録しています。[37] [38]
アンジュアン島とグランドコモロ諸島は、シーラカンスにとって理想的な水中洞窟の生息地を提供しています。これらの島々の海底火山斜面は、急峻に侵食され砂に覆われており、洞窟や岩の裂け目が発達しており、シーラカンスは日中に休息することができます。これらの島々は、シーラカンスの個体群維持に寄与する多くの底生魚類を支えています。[19] [20]
シーラカンスは日中、水深100~500メートル(330~1,640フィート)の洞窟で休息します。他のシーラカンスはより深い海域に回遊します。[3] [9] :37 より 冷たい水(120メートルまたは390フィート以下)はシーラカンスの代謝コストを削減します。サンゴ礁に向かって漂流し、夜間に摂食することで、重要なエネルギーを節約します。[19]日中に洞窟で休息することで、流れに逆らうために費やされるはずだったエネルギーも節約できます。[20]
発見
| 日付 | 説明 |
|---|---|
| 1938年以前 | 西洋では知られていませんでしたが、南アフリカの原住民はこの魚について知っており、「ゴンベッサ」または「マメ」と呼んでいました。[41] |
| 1938年 | (12月23日) 南アフリカ、イーストロンドンの南西30キロメートルで、現代のシーラカンスが初めて発見されました |
| 1952 | (12月21日)コモロ諸島で科学的に知られる2番目の標本が確認されました。それ以来、グランドコモロ島とアンジュアン島沖で200体以上が捕獲されています。 |
| 1988 | ハンス・フリッケがグランドコモロ沖で |
| 1991 | モザンビーク沖、ケリマネの北東24キロメートル沖で初めてシーラカンスが確認されました。 |
| 1995 | マダガスカル島、トゥリアラの南30キロメートルで初めてシーラカンスが記録されました。 |
| 1997 | (September 18) New species of coelacanth found in Indonesia. |
| 2000 | A group found by divers off Sodwana Bay, iSimangaliso Wetland Park, South Africa. |
| 2001 | A group found off the coast of Kenya. |
| 2003 | First coelacanth caught by fishermen in Tanzania. Within the year, 22 were caught in total. |
| 2004 | Canadian researcher William Sommers captured the largest recorded specimen of coelacanth off the coast of Madagascar.[citation needed] |
| 2007 | (5月19日)インドネシアの漁師、ジャスティヌス・ラハマ氏は、 スラウェシ島沖、ブナケン国立海洋公園付近で、体長1.31メートル(4.30フィート)、体重51キログラム(112 ポンド)のシーラカンスを捕獲しました。シーラカンスは隔離されたプールで17時間生存していました。 [42] |
| 2007年 | (7月15日)ザンジバル島の漁師2人が、体長1.34メートル(4.40フィート)、体重27キログラム(60ポンド)のシーラカンスを捕獲しました。この魚は、タンザニア沖の島の北端で捕獲されました。[43] |
| 2019年 | (11月22日)クワズール・ナタール州南海岸。テクニカルダイバーのビデオ映像には、水深69海里で推定体長1.8メートル、体重100キログラムのシーラカンスが記録されていました。[36] |
南アフリカでの最初の発見
{kind=link}

1938年12月23日、トロール船 ネリン号の船長ヘンドリック・グーセンは、チャルムナ川とンセラ川の間のトロール漁を終え、南アフリカのイースト・ロンドンの港に戻りました。彼はいつものように、イースト・ロンドンの小さな博物館の学芸員である友人のマージョリー・コートネイ=ラティマーに電話をかけ、漁獲物の中に何か興味深いものがないか見てみたいかと尋ね、彼女のために取っておいた奇妙な魚について話しました
南アフリカ水生生物多様性研究所(SAIAB、旧JLBスミス魚類学研究所)のアーカイブに保管されている書簡によると、グーセンはこの魚に損傷を与えないよう多大な努力を払い、乗組員にイースト・ロンドン博物館のために取っておくよう指示した。グーセンは後に、この魚は最初に見たときは鋼鉄のような青色だったが、数時間後に「ネリネ」がイースト・ロンドン港に入港した時には、魚は濃い灰色になっていたと語った
自身の著書のどこにもこの生物の記述を見つけられなかった彼女は、友人のジェームズ・レナード・ブライアリー・スミス教授に連絡を取ろうとしたが、教授はクリスマス休暇で留守だった。魚を保存することができなかった彼女は、しぶしぶ剥製師に送った。スミスが戻ると、彼はすぐにそれがシーラカンスの一種であると分かった。シーラカンスはこれまで化石でしか知られていなかった魚のグループである。スミスはこの新しいシーラカンスを、マージョリー・コートネイ=ラティマーと発見された海域に敬意を表して、 Latimeria chalumnaeと名付けた。2人の発見者はすぐに認知され、この魚は「生きた化石」として知られるようになった。1938年製のシーラカンスは今も南アフリカのイースト・ロンドンにある博物館に展示されている。
しかし、標本は剥製になっていたため、鰓と骨格を検査することができず、本当に同一種であるかどうか疑問が残りました。スミスは2つ目の標本の探索を開始しましたが、それは10年以上かかりました。
コモロ
{kind=link}

{kind=link}

スミスは魚の写真、説明、そして懸賞金を記載したチラシを何千枚も配布しましたが、第二次世界大戦で捜索は中断されました。彼はまた、南アフリカ近海で発見された1938年の標本が、通常の生息地から約1800マイル(2900km)南にあったことも知りませんでした。[44] 100ポンドの懸賞金は、当時の平均的な自給自足の漁師にとっては非常に大きな金額でした。14年後、コモロ諸島で1つの標本が発見されましたが、その魚は地元の人々にとって見慣れたものではありません。コモロ諸島のアンジュアン島のドモニ港では、コモロの人々は、漁師が時折誤って捕まえる、ほとんど食べられない魚である「ゴンベッサ」または「マメ」に、これほどの懸賞金が支払われることに困惑していました
2つ目の標本は、1952年のクリスマス直前にコモロの漁師アハマディ・アブダラによって[44]発見されましたが、当初はMalania hunti、後にMalania anjounaeと記述されました。これは、スミス教授の要請で南アフリカ空軍のダコタ戦闘機を派遣し、標本を回収した南アフリカ首相ダニエル・フランソワ・マランにちなんで名付けられました。後に、第一背びれの欠損は、当初は重要な問題と考えられていましたが、標本の生後間もない時期に受けた怪我が原因であることが判明しました。マランは熱心な創造論者で、この原始的な生物を初めて見せられたとき、目を輝かせながら「まあ、醜い。私たちもかつてはこんな風に見えたと言うのですか?」と叫びました[45]。スミスによって回収された標本は、彼が勤務していた 南アフリカのグラハムズタウンにあるSAIABに展示されています
3つ目の標本は1953年9月に、4つ目の標本は1954年1月に捕獲されました。[44]コモロ諸島の人々は絶滅危惧種の重要性を認識しており、偶然捕獲されたシーラカンスを深海に戻すプログラムを設立しました。[要出典]
1968年に亡くなったスミスについては、シーラカンスに関する彼の記述は、1956年に初版が出版された『Old Fourlegs 』に掲載されています。彼の著書『Sea Fishes of the Indian Ocean』は、妻マーガレットがイラストを描き、共著したもので、この地域の 魚類学の標準的な参考文献となっています。
1988年、海洋生物学者のハンス・フリッケは、グランドコモロ島の西海岸沖180メートル(590フィート)の自然の生息地で初めてこの種を撮影しました。 [46]
インドネシアで2番目の種
{kind=link}

1997年9月18日、アルナズとマーク・エルドマン夫妻は新婚旅行でインドネシアを旅行中、スラウェシ島のマナド・トゥアで奇妙な魚が市場に出回っているのを目撃しました。[47]マークはそれがゴンベッサ(コモロシーラカンス)だと思いましたが、青ではなく茶色でした。インターネットで彼らの写真を見た専門家が、その重要性に気づきました。その後、エルドマン夫妻は地元の漁師に連絡を取り、今後この魚を捕獲したら持ってきてくれるよう依頼しました。
1998年7月30日には、体長1.2メートル、体重29キログラムの2匹目のインドネシア産標本が生きたまま捕獲されました。 [40]この標本は6時間生きており、科学者たちはその体色、ひれの動き、そして一般的な行動を写真で記録することができました。この標本は保存され、インドネシア科学研究所傘下のボゴール動物学博物館に寄贈されました。[47]
DNA鑑定の結果、この標本はコモロ諸島の個体群とは遺伝的に異なることが明らかになった。[48] [49]インドネシア産のシーラカンスは、現地ではラジャ・ラウト(海の王)と呼ばれ、表面的にはコモロ諸島で見られるものと同じように見えるが、皮膚の背景色が青みがかった色ではなく、茶色がかった灰色をしている。この魚は、1999年にパリ科学アカデミー紀要(Comptes Rendus de l'Académie des sciences Paris)でPouyaudらによって記載され、学名は Latimeria menadoensisとされた。[50]最近の分子生物学的研究では、2種のシーラカンスの分岐時期は4000万~3000万年前と推定されている。[51]
2007年5月19日、インドネシアの漁師、ジャスティヌス・ラハマ氏は、スラウェシ島北部のマナド沖、ブナケン国立海洋公園の近くで、体長1.3メートル、体重50キログラムのシーラカンスを捕獲しました。30分間水から出た後、まだ生きていたシーラカンスは、海辺のレストラン前の網で囲まれたプールに入れられました。シーラカンスは17時間生き延びました。シーラカンスは通常、水深200~1000メートルに生息しています。シーラカンスは地元当局によって1メートルの深さのプールで泳ぐ様子が撮影され、死亡した後に冷凍されました。AFP通信によると、フランス、日本、インドネシアの科学者がフランス開発研究機構と協力してシーラカンスの解剖を行い、その後遺伝子分析を行う予定です。現在、地元の大学が死骸を研究しています。[42] [52]
南アフリカのイシマンガリソ湿地公園
南アフリカでは、何年にもわたって断続的に捜索が続けられていました。46歳のダイバー、リーハン・バウワーは、1998年6月にシーラカンスを探して命を落としました。
2000年10月28日、モザンビーク国境のすぐ南、セントルシア海洋保護区のソドワナ湾で、ピーター・ベンター、ピーター・ティム、エティエンヌ・ル・ルーの3人の深海ダイバーが104メートルまで潜り、思いがけずシーラカンスを発見しました
「SAシーラカンス探検隊2000」と名乗る一行は、写真撮影機材と数名の追加メンバーと共に帰還しました。11月27日、前日の最初の潜水が失敗に終わった後、ピーター・ベンター、ギルバート・ガン、クリスト・セルフォンテイン、デニス・ハーディングの4名が3頭のシーラカンスを発見しました。最大のものは体長1.5~1.8メートル、他の2頭は1.0~1.2メートルでした。シーラカンスは頭を下にして泳ぎ、洞窟の棚から餌を食べているようでした。一行はシーラカンスのビデオ映像と写真を持ち帰りました
しかし、潜水中にセルフォンテインは意識を失い、34歳のデニス・ハーディングも制御不能な浮上により彼と共に水面に浮上しました。ハーディングは首の痛みを訴え、ボート上で脳塞栓症で死亡しました。セルフォンテインは減圧症の治療のために水中に運ばれた後、回復しました。
2002年3月から4月にかけて、ジェイゴ潜水艇とフリッケ潜水隊はソドワナ沖の深海に潜り、15頭のシーラカンスを観察しました。組織サンプルを採取するためにダートプローブが使用されました。
タンザニア
シーラカンスは2004年以来タンザニアの海岸で捕獲されている。 2003年8月、インド洋沖の小さな島、ソンゴ・ムナラ沖で2匹のシーラカンスが最初に捕獲されたと報告された。その後5か月の間に、体重25~80kgのこの極めて珍しい魚が19匹以上、大量に網にかかったと報告され、2005年1月にも別の1匹が捕獲された。2006年には、オブザーバー紙が、体重110kgものシーラカンスがいると報じた。
シーラカンスの保護のための長期戦略を持つタンガ沿岸域保全開発プログラムの職員は、トロール漁業、特に 日本の船舶によるシーラカンスの生息地付近での捕獲のタイミングと関連性があると考えています。トロール船が網を投げてから数日以内に、サメを狙った深海の刺し網にシーラカンスが見つかったからです。タンザニア沖でのシーラカンスの突然の出現は、無差別なトロール漁法と生息地の破壊によるシーラカンスの個体数への被害により、その将来について深刻な懸念を引き起こしています。[53]
プログラムコーディネーターのハッサン・コロンボ氏は、「トロール船がなければ、シーラカンスは捕獲されない。それだけだ」と述べた。同僚のソロモン・マコロウェカ氏は、タンザニア政府に対し、トロール船の活動を制限するよう圧力をかけてきたと述べた。「トロール船のおかげで、この驚くべきユニークな魚の個体群が発見されたので、感謝すべきだと思います。しかし、これらの貴重なものを破壊する可能性を懸念しています。政府には、トロール船の活動を制限し、シーラカンスについてより深く学び、保護するための適切な研究プログラムへの資金提供をしてほしいと考えています。」[53]
2008年3月の報告書[54]で 、地元の環境NGOであるタンザニア天然資源フォーラムは、ムワンバニ湾で提案されている港湾プロジェクトが、沿岸のシーラカンスの個体群を脅かす可能性があると警告した。[55]
人間との関係
シーラカンスは、人間やおそらく他のほとんどの魚食動物にとって、食料源としては貧弱だと考えられています。シーラカンスの肉には、多量の油、尿素、ワックスエステル、その他の化合物が含まれており、肉に独特の不快な風味を与え、消化を困難にし、下痢を引き起こす可能性があります。鱗自体が粘液を分泌し、それが体から分泌される過剰な油と混ざり合って、シーラカンスはぬるぬるした食べ物になります。[56]シーラカンスがより一般的な地域では、地元の漁師は、シーラカンスが消費者を病気にする可能性があるため、シーラカンスを避けています。[57]その結果、シーラカンスは博物館や個人収集家が切望する以外に、商業的な価値はほとんどありません。[58]
保全
シーラカンスについてはほとんど知られていないため、保全状況の特徴づけは困難です。Frickeら(1995)によると、この種を保護することは重要です。1988年から1994年にかけて、Frickeは1回の潜水で約60頭のL. chalumnaeを数えました。1995年にはその数は40頭に減少しました。これは自然な個体群変動の結果である可能性もありますが、乱獲の結果である可能性もあります。IUCNは現在、L. chalumnaeを「絶滅危惧種」に分類しており[59]、個体数は500頭以下です[9] 。L . menadoensisは、個体数がかなり大きく(10,000頭未満)、危急種とされています[60] 。
シーラカンスに対する主な脅威は、漁業、特に商業的な深海トロール漁業による偶発的な捕獲(混獲)です[61] [62]シーラカンスは、地元の漁師がアブラフィッシュを漁っているときに漁獲されることが多い。漁師は伝統的に夜に漁をするため、アブラフィッシュ(およびシーラカンス)が餌をとる時間帯に漁をするため、アブラフィッシュではなくシーラカンスを釣り上げることがある。科学者がシーラカンスに興味を持つ以前は、捕獲されたシーラカンスは水中に戻されていた。現在では、その重要性が認識され、漁師は科学者や他の当局にシーラカンスを交換している。1980年代以前は、これはシーラカンスの個体数にとって問題だった。1980年代には、国際援助によって地元の漁師にグラスファイバー製の船が支給され、シーラカンスの生息域外、より生産性の高い海域での漁業が行われた。それ以来、船のほとんどのモーターが故障したため、漁師はシーラカンスの生息域に戻らざるを得なくなり、この種は再び危険にさらされている。[9] : 40 [63]
シーラカンスの漁獲量を最小限に抑える方法としては、漁師を海岸から遠ざけること、様々な下剤やマラリア軟膏を用いてアブラフィッシュの需要を減らすこと、博物館でシーラカンスの模型を用いて生きた標本を模倣すること、そして保全の必要性に対する意識を高めることなどが挙げられます。1987年、シーラカンス保全協議会(CCC)はシーラカンスの保全を提唱しました。CCCはコモロ、南アフリカ、カナダ、イギリス、アメリカ、日本、ドイツに支部を置いています。これらの機関は、シーラカンスの保護と個体数増加を促進するために設立されました。[9] : 40 [64]
2014年に「ディープリリースキット」が開発され、民間主導で配布されました。これは、漁師が誤って捕獲したシーラカンスを深海に戻し、海底に着いたフックを外すことを可能にする重り付きのフックアセンブリで構成されています。この方法の有効性に関する決定的な報告はまだ保留中です。[65] [要更新]
2002年、シーラカンスの研究と保全を促進するために、南アフリカシーラカンス保全・ゲノム資源プログラムが開始されました。このプログラムは、生物多様性の保全、進化生物学、能力開発、そして国民の理解に焦点を当てています。南アフリカ政府は、このプログラムに1000万ランドを費やすことを約束しました。[66] [67] 2011年には、シーラカンスを含む海洋動物の生物多様性を保全するために、タンガシーラカンス海洋公園の計画が策定されました。この公園は、生息地の破壊を減らし、絶滅危惧種の餌の入手可能性を向上させることを目的として設計されました。[64]
養殖において
シーラカンスの発見は驚くべき性質を持つため、現代美術、工芸、文学においてしばしばインスピレーションの源となっています。少なくとも22か国が切手にシーラカンスを描いており、特にコモロは12種類のシーラカンス切手を発行しています。また、1000コモロ・フラン紙幣と5CF硬貨にもシーラカンスが描かれています。 [68]
現代日本の文化はシーラカンスに興味を持ち、様々なメディアプロジェクトで取り上げています。ポケモンのメディアフランチャイズでは、レリカンスとして知られるポケモンはシーラカンスをモデルにしています。[69] [70]ビデオゲームシリーズ『あつまれ どうぶつの森』では、シーラカンスはプレイヤーが海で釣りをすることで捕まえることができる珍しい魚です。[71] [72]
参考文献
- ^ 「第7部 脊椎動物」。系統的配列による属群名の収集。2016年10月5日時点のオリジナルからアーカイブ。 2016年6月30日閲覧。
- ^ ab Holder, Mark T.; Erdmann, Mark V.; Wilcox, Thomas P.; Caldwell, Roy L.; Hillis, David M. (1999). 「シーラカンスの2つの現生種?」米国 科学アカデミー紀要。96 (22): 12616–20 . Bibcode :1999PNAS...9612616H. doi : 10.1073/pnas.96.22.12616 . JSTOR 49396. PMC 23015. PMID 10535971 引用エラー:名前付き参照「ref3」が異なる内容で複数回定義されています(ヘルプページをご覧ください)。
- ^ abc Butler, Carolyn(2011年3月)「Living Fossil Fish(生きた化石魚)」ナショナルジオグラフィック:86~ 93
- ^ Musick, JA(2000年)「Latimeria chalumnae(ラティメリア・カルムナエ )」IUCNレッドリスト絶滅危惧種. 2000 e.T11375A3274618. doi : 10.2305/IUCN.UK.2000.RLTS.T11375A3274618.en
- ^ 「シーラカンス、シーラカンスの写真、シーラカンスの豆知識 - ナショナルジオグラフィック」ナショナルジオグラフィック。2011年5月10日。2010年1月14日時点のオリジナルよりアーカイブ。2015年10月28日閲覧。
- ^ 「シーラカンスは研究者の予想より5倍長く、1世紀近く生きる可能性がある」ユーレカレルト。2021年6月17日。2021年6月17日閲覧
- ^ Mahé, Kélig; Ernande, Bruno; Herbin, Marc (2021年6月17日). 「新たな鱗の分析により、100歳を超えるアフリカのシーラカンスの存在が明らかに」. Current Biology . 31 (16). Cell Press : 3621–3628.e4. Bibcode : 2021CBio...31E3621M. doi : 10.1016/j.cub.2021.05.054 . ISSN 0960-9822. PMID 34143958
- ^ 横山正三; 張 煥; ラドルウィマー F. ベルンハルト; ブロウ ネイサン S. (1999). 「コモロ産シーラカンス(Latimeria chalumnae)の色覚の適応的進化」.米国科学アカデミー紀要. 96 (11): 6279–84 . Bibcode :1999PNAS...96.6279Y. doi : 10.1073/pnas.96.11.6279 . JSTOR 47861. PMC 26872. PMID 10339578 .
- ^ abcdefghij Forey, Peter L (1998).シーラカンスの歴史. ロンドン: Chapman & Hall. pp. 1, 3, 6 , 13–16 , 19, 27, 32, 35–40. ISBN 978-0-412-78480-4。
- ^ ab Fritzsch, B. (1987). 「シーラカンス Latimeriaの内耳は四肢動物との類似性を持つ」. Nature . 327 (6118): 153–4 . Bibcode :1987Natur.327..153F. doi :10.1038/327153a0. PMID 22567677. S2CID 4307982
- ^ クラック、ジェニファー・A.(2012年6月27日)『ゲインイング・グラウンド 第2版:テトラポッドの起源と進化』インディアナ大学出版局、391ページ。ISBN 978-0-253-00537-3。
- ^ 「シーラカンス ― 形態学的には玉石混交」リーフクエストサメ研究センター
- ^ ブリト, パウロ・M.; ムニエ, フランソワ・J.; クレマン, ガエル; ジェファール=クリヤマ・リンクス, ディディエ (2010). 「化石シーラカンス Axelrodichthys araripensis (Actinistia: Mawsoniidae) の石灰化した肺の組織学的構造」.古生物学. 53 (6): 1281–90 .書誌コード:2010Palgy..53.1281B. doi : 10.1111/j.1475-4983.2010.01015.x .
- ^ スミス, HW (2018年10月24日). 魚から哲学者へ. リポール・クラシク. p. 92. ISBN 978-5-87392-693-0。
- ^ ピーター・フォリー(1997年11月30日)『シーラカンスの歴史』シュプリンガー・サイエンス&ビジネス・メディア、26ページ。ISBN 978-0-412-78480-4。
- ^ 200ページ、サマンサ・ワインバーグ、2006年。『時を捉えた魚:シーラカンスの探究』ハーパーコリンズ出版社、ニューヨーク、ニューヨーク州
- ^ Makapedua, Daisy Monica; Barucca, Marco; Forconi, Mariko; Antonucci, Niki; Bizzaro, Davide; Amici, Adolfo; Carradori, Maria Rita; Olmo, Ettore; Canapa, Adriana (2011年9月). 「インドネシア産シーラカンスLatimeria menadoensisのゲノムサイズ、GCパーセンテージ、5mCレベル」 . Marine Genomics . 4 (3): 167– 172. Bibcode :2011MarGn...4..167M. doi :10.1016/j.margen.2011.04.001. PMID 21867968. 2020年2月18日閲覧。
- ^ ab ハンス・フリッケ、オラフ・ライニッケ、ヘリベルト・ホーファー、ヴェルナー・ナハティガル (1987). 「自然環境におけるシーラカンスLatimeria chalumnaeの移動」Nature . 329 (6137): 331–3 .書誌コード:1987Natur.329..331F. doi :10.1038/329331a0. S2CID 4353395
- ^ abcd Fricke, H.; Plante, R. (1988). 「インド洋グランデコモロ島における現生シーラカンスLatimeria chalumnaeの生息環境要件」自然科学誌. 75 (3): 149–51 .書誌コード: 1988NW.....75..149F. doi : 10.1007/BF00405310. S2CID 39620387
- ^ abcde Fricke, Hans; Schauer, Jürgen; Hissmann, Karen; Kasang, Lutz; Plante, Raphael (1991). 「洞窟におけるシーラカンスLatimeria chalumnaeの集合体:休息地と社会行動に関する初観察」Environmental Biology of Fishes . 30 (3): 281–6 . Bibcode :1991EnvBF..30..281F. doi :10.1007/BF02028843. S2CID 35672220.
- ^ Bruton, Michael (1991).シーラカンスLatimeria chalumnaeの生態と保全. オランダ:Kluwer Academic Publishers. pp. 313– 339
- ^ ヒスマン、カレン;フリッケ、ハンス;シャウアー、ユルゲン (2008). 「シーラカンス(Latimeria chalumnae)の個体群モニタリング」. Conservation Biology . 12 (4): 759–65 . doi :10.1111/j.1523-1739.1998.97060.x. JSTOR 2387536. S2CID 83504862
- ^ ab Lavett Smith, C.; Rand, Charles S.; Schaeffer, Bobb; Atz, James W. (1975). 「生きたシーラカンス、ラティメリアは卵胎生である」. Science . 190 (4219): 1105–6 . Bibcode :1975Sci...190.1105L. doi :10.1126/science.190.4219.1105. S2CID 83943031.
- ^ 「シーラカンス:5つの豆知識」. AMNH . 2015年10月28日閲覧
- ^ ラングレー、リズ(2021年6月18日)「これらの原始的な深海魚は100歳まで生き、科学者を驚かせる」ナショナルジオグラフィック。2021年6月24日時点のオリジナルからアーカイブ。 2021年6月19日閲覧。
- ^ ブリッグス、ヘレン(2021年6月17日)「『生きた化石』の魚は最長100年生きるかもしれない」BBCニュース。 2021年6月19日閲覧。
- ^ 「シーラカンスは人間と同じくらい長生きする」エコノミスト。2021年6月19日。2021年8月19日閲覧
- ^ ランパート、カトリン・P.;ブラスマン、カトリン;ヒスマン、カレン;シャウアー、ユルゲン;シュヌラ、ピーター;カロウシ、ザホル・エル;ンガトゥンガ、ベンジャミン・P.;フリッケ、ハンス;シャルトル、マンフレッド (2013).「シーラカンスにおける単独雄の父性」(PDF) . Nature Communications . 4 2488. Bibcode :2013NatCo...4.2488L. doi :10.1038/ncomms3488. PMID 24048316
- ^ トリニョ、パブロ;ソト、マティアス;ペレア、ダニエル(2021年2月25日)「シーラカンス科魚類(肉鰭綱、アクティニス亜綱)の包括的な系統解析とマウソニ科およびラティメリ科の構成に関する考察:新旧の方法論的課題と制約の評価」。 歴史生物学。33 (12): 3423– 3443。書誌コード:2021HBio ... 33.3423T。doi :10.1080/08912963.2020.1867982。ISSN 0891-2963。S2CID 233942585
- ^ カダルスマン;萩ユリア菅葉。ローラン・プヨー。レジス・ホッデ。インタヌルフェミ B. ヒスマヤサリ。エンダン・グナイサ;サントソ・B・ウィディアルト。グラム・アラファト氏。フェルリアナ・ウィディアサリ。デヴィッド・ムイヨ;エマニュエル・パラディ(2020年1月13日)。 「インドネシアのシーラカンスの2つの系統間の1,300万年の分岐」。科学的報告書。10 (1): 192. Bibcode :2020NatSR..10..192K。土井:10.1038/s41598-019-57042-1。PMC 6957673。PMID 31932637。
- ^ ab Amemiya, Chris T.; Alföldi, Jessica; Lee, Alison P.; Fan, Shaohua; Philippe, Hervé; MacCallum, Iain; Braasch, Ingo; Manousaki, Tereza; Schneider, Igor; et al. (2013年4月18日). 「アフリカのシーラカンスのゲノムは四肢動物の進化に関する知見を提供する」. Nature . 496 (7445): 311–6 . Bibcode :2013Natur.496..311A. doi :10.1038/nature12027. PMC 3633110. PMID 23598338 .
- ^ Northcutt, R. Glenn (1986). 「肺魚の神経形質と肉鰭綱の系統発生への影響」(PDF) . Journal of Morphology . 190 (S1): 277– 297. Bibcode :1986JMorp.190S.277N. doi :10.1002/jmor.1051900418. hdl : 2027.42/50281 . S2CID 35473487.
- ^ Stromberg, Joseph. 「DNA配列解析により、シーラカンスは海と陸をつなぐミッシングリンクではなかったことが明らかに」. Smithsonian Magazine
- ^ Naville, Magali; Chalopin, Domitille; Volff, Jean-Nicolas (2014). 「種間挿入多型解析により現生シーラカンスにおける転移因子の最近の活動が明らかに」. PLOS ONE . 9 (12) e114382. Bibcode :2014PLoSO...9k4382N. doi : 10.1371/journal.pone.0114382 . PMC 4255032. PMID 25470617
- ^ 二階堂正人、佐々木健、エマーソンJJ、相原光人、ムジガニ・セムブア・I.、ブデバ・ヨハナ・L.、ンガトゥンガ・ベンジャミン・P.、岩田正光、阿部義孝 (2011年11月1日). 「タンザニア北部沖における遺伝的に異なるシーラカンスの個体群」.米国科学アカデミー紀要. 108 (44): 18009–18013 .書誌コード:2011PNAS..10818009N. doi : 10.1073/ pnas.1115675108 . PMC 3207662. PMID 22025696
- ^ ab フレイザー、マイケル・D.;ヘンダーソン、ブルース・AS;カーステンス、ピーター・B.;フレイザー、アラン・D.;ヘンダーソン、ベンジャミン・S.;デュークス、マーク・D.;ブルトン、マイケル・N.(2020年3月26日)「南アフリカ、クワズール・ナタール州南海岸沖で生きたシーラカンスが発見される」。南アフリカ科学ジャーナル116(2020年3月/4月号)。doi :10.17159 /sajs.2020/ 7806
- ^ Augy Syaihailatua (2015年3月30日). 「インドネシア海域での生きた化石探し」The Conversation .
- ^ リック・ヌレンズ、ルーシー・スコット、マーク・エルビン(2011年9月22日)「シーラカンス(ラティメリア属)の既知の全標本の最新目録」(PDF)。スミシアナ:2。 2018年8月18日時点のオリジナル(PDF)からのアーカイブ。
- ^ イジコ南アフリカ博物館、ケープタウン
- ^ ジョセフ・S・ネルソン著(2006年)『世界の魚たち』。ジョン・ワイリー・アンド・サンズ社ISBN 0-471-25031-7
- ^ 噂か現実か:未確認動物学の生き物、リスト7位。Livescience.com
- ^ ロイター通信(2007年)「インドネシアの漁師が古代の魚を捕獲」、ロイターUK、2007年5月21日、2007年7月16日閲覧
- ^ ロイター (2007)、「ザンジバルの漁師、古代の魚を陸揚げ」、ロイターUK、2007年7月15日、2009年12月13日閲覧。
- ^ abc レイ、ウィリー (1965年10月)。「銀河の15年 — FYIの13年」『For Your Information』、ギャラクシーサイエンスフィクション、 84~ 94ページ。
- ^ ワインバーグ、サマンサ、2006年、『時を捉えた魚:シーラカンスの探究』、73ページ。ハーパーコリンズ出版社、ニューヨーク、ニューヨーク州。
- ^ フリッケ、ハンス (1988年6月)。「シーラカンス:時が忘れた魚」、ナショナルジオグラフィック、173 (6): 824~ 828
- ^ ab Jewett, Susan L., 「生きた化石シーラカンスの足跡を辿って」ワシントン・ポスト、1998年11月11日、2007年6月19日閲覧。
- ^ Erdmann, Mark V. (1999年4月). 「インドネシア海域で科学者に知られる最初の生きたシーラカンスに関する報告」Environmental Biology of Fishes . 54 (#4). Springer Netherlands: 439– 443. Bibcode :1999EnvBF..54..439E. doi :10.1023/A:1007584227315. S2CID 46211870. 0378-1909 (印刷版) 1573-5133 (オンライン版).
- ^ ホルダー、マーク・T.、マーク・V・エルドマン、トーマス・P・ウィルコックス、ロイ・L・コールドウェル、デイビッド・M・ヒリス (1999). 「シーラカンスの2つの現生種?」米国科学アカデミー紀要. 96 (22): 12616– 12620. Bibcode :1999PNAS...9612616H. doi : 10.1073/pnas.96.22.12616 . PMC 23015. PMID 10535971.
- ^ Pouyaud, L.; S. Wirjoatmodjo; I. Rachmatika; A. Tjakrawidjaja; R. Hadiaty & W. Hadie (1999). 「シーラカンスの新種:遺伝学的・形態学的研究」.アカデミー科学誌, 第3シリーズ. 322 (4): 261– 267. Bibcode :1999CRASG.322..261P. doi :10.1016/S0764-4469(99)80061-4. PMID 10216801.
- ^ Inoue JG; M. Miya; B. Venkatesh; M. Nishida (2005). 「インドネシア産シーラカンスLatimeria menadoensis(肉鰭綱:シーラカンス目)のミトコンドリアゲノムと2種のシーラカンス間の分岐時期の推定」Gene . 349 : 227–235 . doi :10.1016/j.gene.2005.01.008. PMID 15777665.
- ^ 「古代インドネシアの魚は『生きた化石』」 Wayback Machineに2008年12月5日アーカイブ、Cosmos Online、2007年7月29日
- ^ 「深海トロール船によって絶滅の危機に瀕した恐竜の魚」オブザーバー、2006年1月8日、2007年6月18日閲覧。
- ^ 「タンガ島はムワンバニ湾に新しい港を必要としているか?」Wayback Machineに2010年9月12日アーカイブ、タンザニア天然資源フォーラム、2008年3月5日、2009年2月25日閲覧。
- ^ 「先史時代の深海シーラカンスの個体数は恐竜と同じ道を辿るかもしれない」mongabay.com、2009年2月25日、2009年2月25日閲覧
- ^ 「クリーチャー特集:シーラカンスに関する10の楽しい事実」Wired . 2015年3月2日. 2015年10月30日閲覧.
- ^ アダムス、セシル(2011年12月30日).「絶滅危惧種の先史時代の魚の美味しいレシピを知っていますか?さらに:カリブーはアラスカの石油パイプラインが好きですか?」The Straight Dope .
- ^ パイパー、ロス(2007年)『Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals』Greenwood Press、249ページ。ISBN 978-0-313-33922-6。
- ^ Musick, JA (2000). 「Latimeria chalumnae」. IUCNレッドリスト絶滅危惧種. 2000 e.T11375A3274618. doi : 10.2305/IUCN.UK.2000.RLTS.T11375A3274618.en . 2021年11月13日閲覧
- ^ Erdmann, M. (2008). 「Latimeria menadoensis」. IUCNレッドリスト絶滅危惧種. 2008 e.T135484A4129545. doi : 10.2305/IUCN.UK.2008.RLTS.T135484A4129545.en . 2021年11月13日閲覧.
- ^ Gilmore, Inigo (2006年1月7日). 「深海トロール船によって絶滅の危機に瀕した恐竜の魚」.オブザーバー.
- ^ 「シーラカンス」.アニマルプラネット. 2012年8月27日. 2015年10月29日閲覧
- ^ フリッケ、ハンス、ヒスマン、カレン、シャウアー、ラファエル・プランテ (1995). 「シーラカンスにとってさらなる危険」. Nature . 374 (6520): 314–5 .書誌コード:1995Natur.374..314F. doi : 10.1038/374314a0 . S2CID 4282105.
- ^ ab 「シーラカンス(Latimeria chalumnae)に関する絶滅危惧種保護法現状レビュー報告書」(PDF) . nmfs.noaa.gov . オリジナル(PDF)から2015年9月7日にアーカイブ。2015年10月30日閲覧
- ^ 「タンザニアのアフリカシーラカンスDPSを絶滅危惧種法に基づき絶滅危惧種に指定するための提案規則」NOOA(米国) 。 2015年10月30日閲覧。
- ^ 「南アフリカ、シーラカンス・プログラムの計画を発表」(プレスリリース)。Science in Africa。2002年2月。2015年4月2日時点のオリジナルよりアーカイブ。 2013年4月19日閲覧。
- ^ 「南アフリカのシーラカンス保全およびゲノム資源プログラム」。アフリカ保全財団。2015年4月2日時点のオリジナルよりアーカイブ。 2013年4月19日閲覧。
- ^ Smith, JLB (2017). The Annotated Old Four legs . Cape Town: Struik Travel & Heritage. pp. 322– 327. ISBN 978-1-77584-501-0 OCLC 1100871937
- ^ メンデス、アウグスト・B.、ギマランイス、フェリペ・V.、エイラード=シルバ、クララ・BP、シルバ、エドソン・P. (2017).「ポケモンの魚類学的多様性」.ジャーナル・オブ・ギーク・スタディーズ. 4 (1): 39– 67. ISSN 2359-3024 . 2019年7月12日閲覧
- ^ Williams, Leoma (2023年5月24日). 「本物の野生動物にインスパイアされたポケモンキャラクター20選」BBC Wildlife . 2023年12月10日閲覧。
- ^ 「あつまれ どうぶつの森」で最も捕まえにくい魚、実生活では奇妙な裏話が隠されている。ポピュラーサイエンス誌。2020年4月8日。2023年12月10日閲覧。
- ^ ネアン、レイシー(2020年4月1日)「あつまれ どうぶつの森:ゲームで最もレアな魚を捕まえる方法」。コミックブックリソース。 2023年12月10日閲覧。
