| マイムーラペルタ 時代範囲:ジュラ紀後期、
| |
|---|---|

| |
| ワイオミング恐竜センターの復元骨格 | |
| 科学的分類 | |
| 界: | 動物界 |
| 門: | 脊索動物門 |
| 綱 | 爬虫類 |
| クレード | 恐竜 |
| クレード | †鳥盤類 |
| クレード | †甲羅目 |
| クレード | †角竜類 |
| クレード | †真角竜類 |
| 科: | †ノドサウルス科 |
| 属: | †ミムーラペルタ カークランド・カーペンター, 1994 |
| タイプ種 | |
| †ミムーラペルタ・マイシ | |
マイムーラペルタ(化石が最初に採集されたマイガット・ムーア採石場の発見者の名前とギリシャ語の「盾」を意味するπέλταを組み合わせたものに由来)は、アメリカ合衆国コロラド州西部とユタ州中央部にまたがるジュラ紀後期(キンメリッジアン-ティトニアン、約1億5500万年前から1億5000万年前)のモリソン層(ブラッシー盆地層)にていたノドサウルス 科の曲竜類である。この動物はMymoorapelta maysiという1種のみが知られており、標本はわずかしか知られていない。最も完全な標本はマイガット・ムーア採石場のホロタイプ個体で、皮骨、部分的な頭蓋骨、椎骨、その他の骨が含まれている。 1994年にジェームズ・カークランドとケネス・カーペンターによって初めて記載されました。ガーゴイレオサウルスとともに、最も古いノドサウルス科の1つです。
ミムーラペルタは既知のノドサウルス科の中では小型の部類に入り、最大の標本でも体長は3メートル(9.8フィート)と推定されている。吻部は狭く、背面から見ると頭骨はほぼ三角形で、額から後方に伸びた2本の大きな角と、その下から頬骨に向かって後方下方に伸びた2本の角があった。ミムーラペルタでは5種類の装甲が確認されており、体側面の細長く鋭い棘から、骨盤上部を覆う、耳小骨と呼ばれる小さな皮骨でできた巨大な仙骨の盾まで様々である。棍棒状の尾を持つアンキロサウルス科とは対照的に、尾にはミムーラペルタが防御に使用したと思われる 棘があった。
ミムーラペルタはモリソン生態系において低地の草食動物であり、ソテツや針葉樹を食べていました。これは、同じ採石場から発見された高地の草食動物であるアパトサウルスとは対照的です。大型獣脚類の アロサウルスやケラトサウルス、そしておそらく鳥盤類の ナノサウルスを含む他の恐竜のグループも生息していました。これらの獣脚類はいずれもミムーラペルタと共存しており、前者がミムーラペルタを捕食していたという証拠があります。
発見と命名

オリジナルの発見
マイガット・ムーア採石場は、1981年3月、コロラド州中西部メサ郡で、ハイカーのヴァネッタ・ムーアとピート・マイガット、マリリン・マイガット夫妻によって初めて発見されました。採石場の地層は、上部ジュラ紀のモリソン層のブラッシー・ベイスン層に由来しています。[1] [2]マイムーラペルタの化石は1990年に初めてこの採石場で発見され、それ以来、150平方メートル(1,600平方フィート)の採石場の25メートル(27ヤード)にわたって160以上の化石が散在して発見されています。[3] [1]これらの要素のいくつかは単一の成体個体の一部であり、1994年にアメリカの古生物学者ジェームズ・カークランドとケネス・カーペンターによって記載されました。この標本(MWC 1815)は、この種のホロタイプに指定され、保存された噛み跡を持つ左腸骨(寛骨)が含まれています。[1]カークランドとカーペンターはこの種をMymoorapelta maysiと命名しました。属名は、採石場を発見したマリリン・マイガットとムーア夫妻の名前と、保存された装甲にちなんで「盾」を意味するギリシャ語の語根peltaに由来しています。 [1]種小名は、マイガット・ムーア採石場の最初の発掘に資金を提供した、ダイナメーション・インターナショナル・コーポレーション・アンド・ソサエティの会長、クリス・メイズにちなんで付けられました。[1] [3]ミムーラペルタは北アメリカで命名された最初のジュラ紀の曲竜であり、近縁のガルゴイレオサウルスは4年後の1998年に命名されました。[4] [1]採石場から発掘されたタイプ標本のその他の部分には、いくつかの背椎と尾椎、肋骨、孤立した四肢骨、および基底的曲竜の特徴的な三角形の頸椎を含む多くの皮骨が含まれています。[5] [3] [1]採石場で発見され、ミムーラペルタと呼ばれるいくつかの皮骨は、より高次の地層から来ており、2番目の個体が保存されていることを示唆しています。[3]
後の発見
.png/1280px-Holotype_left_ilium_of_Mymoorapelta_maysii_(MWC_1815).png)
1998年、コロラド州カクタスパークでミムーラペルタとされる部分的な骨格がカークランドらによって記載されたが、当時はまだ化石の準備段階にあった。[5]骨格には仙骨、尾椎、シェブロン(恐竜の尾椎の下側に付いている骨)、足、骨盤の上部を覆う装甲の一種である仙骨盾の一部を含む多くの皮膚装甲の自然な型と体の化石が保存されていた。頭骨の素材が保存されていると言及されていたが、まだ準備段階で記載されていない。[6] [1]足と仙骨盾の素材の一部は関節で保存されており、この分類群の関節標本として知られているのはこれが唯一である。[5] 2010年の論文要旨で、カークランドらは、マイガット・ムーア採石場で、ほぼ完全な頭蓋骨と恥骨と大腿骨を除く頭蓋骨以降のすべての要素を含む、ミムーラペルタのより多くの要素が発見されたと述べています。 [5] [7] [3]マイガット・ムーア採石場とカクタスパークで発見された化石はすべて、現在フルータにある西コロラド州の恐竜ジャーニー博物館に保管されています。[3] [1]
2015年には、ケイティ・トレメインらによる論文抄録が発表され、ユタ州ハンクスビル近郊のハンクスビル・バーピー採石場で発掘されたミムーラペルタの新標本について言及されている。[8]この標本には、24枚の皮骨、肋骨3本、椎骨1個、大腿骨1個が保存された個体が含まれているが、まだ発掘されていない標本が多数ある。 2016年には、ニューメキシコ州アルバカーキ郊外のピーターソン採石場で採掘された背側の皮骨1個の発見が報告されている。[9]この皮骨は形態学的には(外部解剖に基づく)ミムーラペルタに類似しているが、地層学的には(岩石層に基づく)ガーゴイレオサウルスに近い。[9]
説明

ミムーラペルタは、モリソン層で発見された最も小型のアンキロサウルス類の一つであり、四足歩行恐竜としても知られている。しかし、個体数は少なく、既知の化石のうち詳細な記載がされているのはごく一部に過ぎない。カクタスパーク標本は、1998年に全長約3メートル(9.8フィート)と推定された、これまでに発見された個体の中で最大のものである。 [5]タイプ標本に基づく推定体重は、約300~562キログラム(661~1,239ポンド)である。[10] [6] [11]
頭骨と歯列

頭骨については特に詳細な説明はされていないが、保存された形態(外部解剖学)は、同じくモリソン層から産出するガルゴイレオサウルスのものと非常によく似ている。 [7]ガルゴイレオサウルスは吻が狭く、背面(上面)から見ると頭骨はほぼ三角形で、額の後ろから後方に伸びた2本の大きな角と、その下の頬骨から後方下方に伸びた2本の角があった。1998年に記載された保存された頬骨はくさび形で装甲形態をしており、アンキロサウルス類のように腹外側部(下方および外方)に突き出ている。[5]前上顎骨(吻端の骨)は、ガルゴイレオサウルスのように狭く歯があり台形の鼻先を持っていたと考えられる。[12]頭骨後部では、方形骨関節部(頭骨と下顎の接触部)がわずかに内側(体の正中線に向かって)を向いており、前後方向(前から後ろ)にわずかに長くなっている。方形骨の遠位面、特に内側顆は頑丈で前後方向に長くなっている。すべてのアンキロサウルス類において、顎関節(頭骨と顎骨を繋ぐ関節)は咬合面(歯の接触面)よりも腹側に位置していた。ノドサウルス類では、冠状突起(下顎の骨の突起)が非常によく発達しており、顎関節が大きく陥没しているため、相対的に咬合力が高くなっていた。
ほとんどのアンキロサウルス類は異歯性歯列(歯の形態における変異)を示すが、ガルゴイレオサウルスにはそれが見られず、前上顎歯、上顎歯、歯骨歯においてわずかな形態的差異しか見られない。アンキロサウルス類の歯は唇舌方向に圧縮され、葉状(葉形)を呈し、先端咬頭と近心縁および遠心縁に沿った二次咬頭を有する。ノドサウルス類の歯は刃状で、大きく、通常アンキロサウルス類の基本咬頭よりも複雑であり、歯冠が大きく、エナメル質表面が粗く、縁咬頭の溝と合流する溝、そして狭窄した帯状部(歯冠基部を囲む棚状の隆起)を有する。アンキロサウルスの歯の摩耗面は大きく異なるが、ノドサウルス科ではアンキロサウルス科よりも摩耗面が広く、傾斜が急であるのが一般的で、形状に制約された機能の根本的なパターンを示している。ガルゴイレオサウルスやガストニアのような初期のノドサウルス科では、翼状骨(口蓋の皮膚の一対の骨)が前後方向および内外方向にまだ十分に発達していないが、後の種では翼状骨がはるかに幅広で、側翼がより頑丈になり、基底型よりも前後方向に拡大して遠位方形顆の高さに達している。これは、後のノドサウルス科ではアンキロサウルス科よりも翼状筋が発達していたことを示唆しており、ノドサウルス科の顎内転筋システムの方が効率的であったことを反映している。ノドサウルス類の下顎内転筋室と冠状突起は、アンキロサウルス類よりも一般的に高く、筋肉の大きさと付着部の違いを示唆している。[12]
頭蓋骨より

ミムーラペルタの頭蓋後部(頭蓋骨の後ろ)の骨格の大部分が知られている。例えば、多くの頸椎(首の椎骨)が知られているが、その多くは十分に調査・発掘されていない。頸椎では、椎体の面(端)は高さよりも幅が広く、ハート型の形態を呈している。椎体全体は前後方向に短く、長さは幅とほぼ同じである。神経棘は短く太く、円形の神経管を有している。後椎弓突起(椎弓の後方突出部)の間には、深い涙型の窩(骨の小さな開口部)が存在する。[5]ミムーラペルタからはいくつかの背椎(背中、胸椎と呼ばれることもある)が知られており、白亜紀のノドサウルス類の椎骨と比較すると独特である。椎体は円筒形で、後方に長く、サウロペルタやドラコペルタのような後のノドサウルス科の他のものよりも側方への圧縮が少ない。前背椎は、前椎弓突起(椎弓の前方突出部)の間に三角形の窪みがある点で独特であり、前椎弓突起も椎骨の前端では非常に長くなっている。一方、後椎弓突起は後端が切断されており、左右の後椎弓突起は大きな溝で分けられている。中央の背椎は、腹側が丸く、底部に弱いキールがある点で異なっている。背椎の横突起は急激に上昇せず、代わりに基底的曲竜の状態に似て徐々に高くなるが、ノドサウルス科のものと異なる。ミムーラペルタは13本の肋骨を持ち、他のアンキロサウルス類とは対照的に、全てが三角形の断面と弱いT字型を呈している。また、ミムーラペルタは4本の仙肋骨を持つ数少ないアンキロサウルス類の一つであり、この特徴を共有するのは他に3つの分類群のみである。[5] [13]

ミムーラペルタをはじめとするノドサウルス類の尾椎は、幅よりも長く、背腹方向(上下方向)に扁平化しており、前尾椎に比べて神経棘が縮小している。血棘は丸みを帯びた手斧のような形状をしている。前接合骨は短く、椎体前縁からわずか25%以下しか突出していない。この特徴はノドサウルス類と基底アンクロサウルス類にのみ見られるが、リャオニンゴサウルスやより派生したアンキロサウルス類のような分類群では、前接合骨が少なくとも全長の50%にわたって重なり、より癒合している。ミムーラペルタは他のノドサウルス類のような尾棍の柄や突起を持たず、そのため強直が比較的少ない。尾椎の前部から後部への移行は、中尾椎で急激な移行があるアンキロサウルス科に比べるとそれほど急激ではない。この急激な移行がないため、尾に柔軟性が与えられ、これはアンキロサウルス科の強く癒合した尾と比較するとノドサウルス科に独特である。[14]椎体と神経棘はガルゴイレオサウルスと比較するとはるかに短い。[5] ミムーラペルタは、他の恐竜と同様に、尾椎の下側に沿ってV字型構造を保存しており、他の多くのアンキロサウルス類と同様に、V字型構造は椎体の下後部に強く癒合し、血弓は細長い椎体と平行に伸びている。これらの弓状骨は後方よりも前方に大きく伸びており、血管管の下に連続した骨構造を形成しています。保存されているV字型骨の一つは長さ9.1センチメートル(3.6インチ)で、後方に強く反り返っています。[1] [5]
ミムーラペルタの肩甲骨棘は、他のアンキロサウルス類や、近くのドライ・メサ採石場で発見された不確定なアンキロサウルスの肩甲骨と同様に、関節窩の反対側に垂直な隆起を形成している。骨盤部では、ミムーラペルタは基盤的なアンキロサウルス類とより派生したアンキロサウルス類のいくつかの特徴の中間的な特徴を示している。坐骨は他のノドサウルス類やポラカントゥス類のように曲がっており、まっすぐな坐骨はアンキロサウルス類の派生した特徴であることがわかる。前肢では、尺骨に幅広く大きな肘頭突起があり、しわが保存されている。冠状突起は尺骨の幹の約3分の1のところで発達するが、ステゴペルタやポラカントゥスなどの分類群では4分の1のところで発達する。この属は、脚において、エドモントニアなどの他のノドサウルス類とは対照的に、長く短い指骨を持つ幅広い中足骨を保存しており、側面から見ると末節骨は三角形で尖っている。[1] [5]
装甲

ミムーラペルタの顕著な特徴はその装甲で、その大部分は体の側面と背面から突き出た大きな棘状の皮骨 (骨質の装甲) で構成されていた。これらは関節部分が発見されていないため、体のどこに付いていたのかは不明だが、他のノドサウルス科やアンキロサウルス類の保存状態の良い骨格に基づいて位置を推測することはできる。ミムーラペルタには、5 種類の異なる装甲が保存されている。大きく中空の基部を持つ細長い棘、狭く非対称な基部を持つ薄い三角形の板、丸くて硬い基部を持つ小さな刃のような棘、独立した平らで竜骨状の鱗板、および 1 枚の装甲板に融合した鱗板である。大きな棘板は首の基部から外側に伸び、先端が後方に曲がっていたと推測されている。[5] [1]ボレアロペルタの証拠も、大きなケラチン鞘によって棘が延長していたことを裏付けている。[15]前述の薄い三角形の板は8枚が知られており、ポラカントゥスの体のさらに下の部分にも同様に保存されている。[16] [1]カークランドとカーペンターは、これらの板のうち最大のものは尾の基部に付着し、体の側面に沿って横方向に突出していたと理論づけた。ミムーラペルタの標本には、長さ11センチメートル(4.3インチ)のものが保存されている。[5] [1]
1994年には、小さな三角形のプレートが1枚発見され、薄い刃のような形態を保っているものの、その位置は不明である。[1]前述のキール付きの平らな甲板のうち12枚は、1994年にミムーラペルタの標本から発見されたが、キール付きの楕円形と、均一で弱い陥凹のある外面、神経血管溝(血流と恒常性をコントロールするシステム)と孔(骨内の小さな開口部)の欠如を保っている。[1]甲板の内部構造は、半分のスペースが海綿骨で構成され、残りの半分は厚い外部皮質と基底皮質で構成されている。[17]
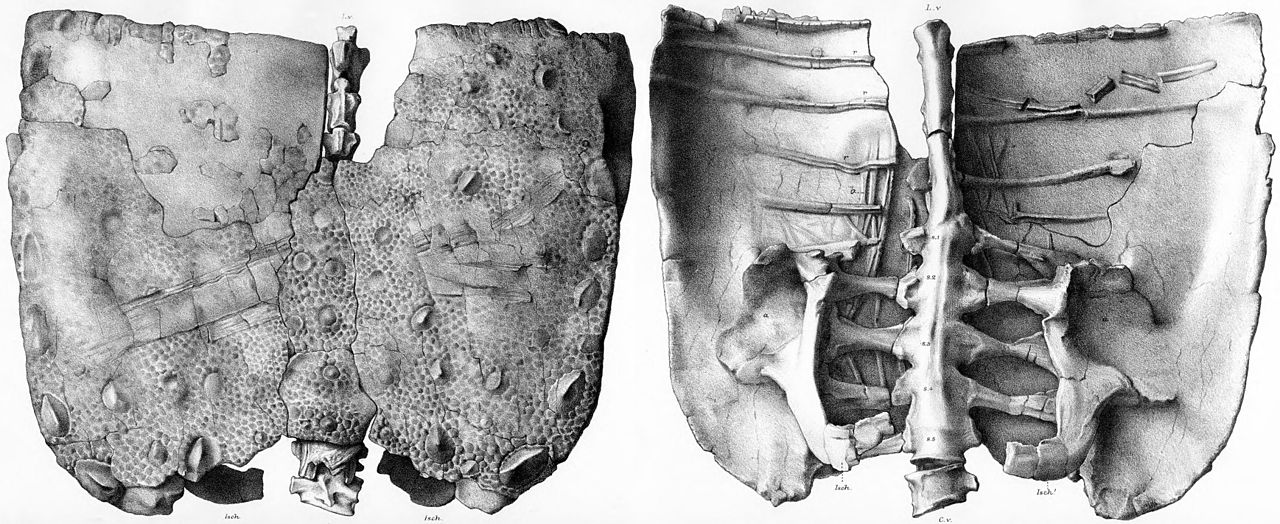
最後に、皮膚装甲の小片にはロゼット(大きな骨皮がより小さな骨皮に囲まれ、花のような配置になっている)が保存されており、中央の大きな鱗板とその周囲を小さな耳小骨が取り囲んでいる。[5] [1]この装甲片は仙骨シールドに由来する。仙骨シールドは、保護のために仙骨と骨盤全体に渡って接続された小さな装甲の大きなプレートである。より完全な仙骨シールドがカクタスパークの標本で保存されており、骨盤の幅が50センチメートル(20インチ)であるため、完全な仙骨シールドの幅は70センチメートル(28インチ)であると推定されている。[5] [1]仙骨シールドには、より大きな鱗板の中央の2列も保存されており、各列には7枚が保存されている。[5] ミムーラペルタはカテゴリー2の仙骨盾を保存しており、これは隆起した突起(キール状の骨皮)と骨化した骨皮のロゼットを特徴としています。このカテゴリーの仙骨盾は主に「ポラカントス科」に見られますが、[18] [19]このグループの地位は一般的に不安定で、この特徴はシャモサウルスなどの一部のアンキロサウルス科にも知られています。[20] [21] [19]カテゴリー2の盾はカテゴリー3の盾に最も似ていますが、後者はロゼットのない多角形の骨皮を保存しています。 [19]しかし、ビクトリア・アーバー らは、このカテゴリー2の盾はカテゴリー3の盾と最も類似していると述べています。(2011) は、カテゴリ 1 の盾はカテゴリ 2 の個体発生 (成長) 段階に過ぎない可能性があると示唆しました。なぜなら、形態が非常に似ているものの、共骨化していないためです。共骨化は、アルマジロやワニなどの現生の装甲動物に存在する個体発生の特徴です。[19] [22] [23]仙骨の盾を明確なカテゴリに分類する現在の方法は、2011 年に Arbour、Michael E. Burns、 Philip J. Currieが発表した論文に端を発し、3 つのカテゴリそれぞれに 1 つずつ、3 つの異なる特徴が示されています。カテゴリ 1 の盾は非骨化皮質、カテゴリ 2 の盾は骨化したロゼット、カテゴリ 3 は均一な大きさの骨化した多角形を特徴とします。[24]
背中と尾の上部の表面は、サウロペルタの推測通り、より大きな装甲板の間に小さな皮骨(耳小骨と呼ばれることもある)で覆われていたが、[25]ミムーラペルタの仙骨の盾以外では耳小骨そのものの存在は確実には分かっていない。[5] [1]ミムーラペルタの装甲はガルゴイレオサウルスの装甲とは異なり、前者は堅固な円錐形の装甲であるのに対し、後者は薄壁の装甲である。[4]
古生物学
食事と摂食

ミムーラペルタは近縁種と同様に低木を食草とする草食動物で、ジュラ紀には複雑なイネ科植物が少なかったため、当時の植物相を支配していたシダ、ソテツ、針葉樹を食べていた可能性が高い。 [26]ミムーラペルタのようなノドサウルス科の動物は鼻先が細く、これは草食動物の幅広い鼻先とは対照的に、今日の選択的ブラウザ動物に見られる適応である。[27] [12]アンキロサウルス類では、ジュラ紀と中期白亜紀に生息した細く尖った鼻先を持つ種が、現生哺乳類のブラウザ動物に類似し、最も選択的な摂食者であったようだ。これは、栄養価の低い食物を大量に摂食するように適応した後期のアンキロサウルス類とは著しい対照をなしている。アンキロサウルス類のピナコサウルスやノドサウルス類のエドモントニアのような分類群では、舌骨が完全に保存されていることから、アンキロサウルスは肉質で筋肉質な舌を持っており、地面近くに生える植物の摂食に役立っていたと考えられます。[28] [12]舌の突出と把持については確実なことは分かっていませんが、キリンのように、舌を使って食物を掴むことは、後期アンキロサウルス類でも行われていた可能性があります。エドモントニアとパノプロサウルスの頬板の保存は、肉質の頬と咀嚼の証拠となります。頬板は歯列を覆うことで防御と、食事中の食物の損失防止の両方の役割を果たしていました。[29]
歯の咬合はミムーラペルタやその近縁種であるガルゴイレオサウルスには直接保存されていないが、白亜紀の他のノドサウルス科動物では発見されている。ノドサウルス科動物の歯の咬合の最も古い証拠はサウロペルタで発見され、力のかかるストロークが垂直方向に動くという基本的な動きが示された。後のノドサウルス科動物とは対照的に、歯と歯の接触は偶発的または局所的であり、ミムーラペルタとガルゴイレオサウルスには二相性の顎の機構が欠けていた。[12]アンキロサウルスは一般に、同時代の白亜紀の角竜類やハドロサウルス科動物 よりも摩耗パターンが多様である。アンキロサウルスは繊維質や木質の植物を食べていなかったかもしれないが、硬い葉や果肉の多い果実など、多様な食生活を送っていた可能性がある。[28]
生息地の好み
ディプロドクス科の竜脚類との競争により、ミムーラペルタは開けた環境に適応していたと考えられる。[30] ミムーラペルタの分布域は非常に限られており、ハンクスビル=バーピーの標本とアルバカーキの標本候補に加えて、コロラド州西部の2か所でのみ発見されている。[1] [5] [8] [9]これは、ミムーラペルタが特定の植物種を好んで食べるため、分布域が限られているためと考えられる。ハンクスビル=バーピーの標本の化石化過程(どのように、どこで分解して化石となったか)は、この個体が内陸で死亡したことを示唆しており、これはこれまでノドサウルス類の沿岸分布と推測されていたこととは対照的である。[8] [31] [32]
四肢の動き
1978年にクームズが行ったアンキロサウルスの前肢の筋肉の復元図は、前肢が動物の体重の大部分を支え、前足に大きな力を伝えるように適応していたことを示唆しています。おそらく食物収集のためでしょう。さらにクームズは、アンキロサウルスは優れた掘削能力を持っていた可能性があると示唆しましたが、蹄のような手の構造は掘削能力を制限していたと考えられます。アンキロサウルスは動きが遅く、鈍重な動物だった可能性が高いですが、[33] [34]必要に応じて素早い動きも可能だった可能性があります。[35] ミムーラペルタは非常に短い四肢とさらに短い遠位四肢要素を保存しており、これは他のノドサウルス科で知られているより長い遠位四肢要素とは対照的ですが、アンキロサウルス科やポラカントゥスに見られる状態と似ています。[7]
装甲と尾

アンキロサウルス類の装甲は防御目的で、主に攻撃してくる肉食動物を撃退するために使用されていたという見解が一致しています。[36]これらの動物の尾には複雑な進化の歴史があり、ミムーラペルタのような基底的な分類群は、両側と上部に尖った皮骨を持つ柔軟な尾を持っています。前述の尾の柔軟性は、後のアンキロサウルス類やノドサウルス類の硬い尾よりも広い可動範囲を可能にしました。[37]尾は捕食者からの防御にも使用されていた可能性があると考えられていますが、2022年のArbourらによるより最近の研究結果では、アンキロサウルス類の尾が種内戦闘に使用され、主にここでの機能を果たすように進化した可能性も検討されており、ミムーラペルタやガルゴイレオサウルスのようなより基底的な分類群でも同様の使用があったことを示唆している可能性があります。[38]
分類
ミムーラペルタはノドサウルス科の最も基底的な属の一つで、ノドサウルス科は絶滅した中型から大型の、がっしりとした体格の四足歩行の草食恐竜で、小さな葉のような歯を持っていました。この系統群は1890年にオスニエル・チャールズ・マーシュによって命名されました。彼は、ノドサウルス属が持つ厚い皮の装甲、硬い骨、大きな前肢、そして有蹄類の足跡に基づいて、白亜紀のノドサウルス属をタイプ属に指定しました。[39] [40]ノドサウルス科はジュラ紀後期(約1億5500万年前)に初めて進化し、ミムーラペルタと同サイズのガーゴイレオサウルスは、コープ則に従って白亜紀まで続く進化の放散の初期のメンバーでした。[41] [4]ノドサウルス類は白亜紀後期(約6600万年前)の白亜紀-古第三紀絶滅イベント中に他の非鳥類恐竜とともに絶滅した。

ノドサウルス科は、近縁のアンキロサウルス科とともに、真正アンキロサウルス類に分類される。ノドサウルス科は、アンキロサウルス類がより最近、前期白亜紀に進化したという点と、いくつかの異なる解剖学的特徴においてアンキロサウルス類と異なる。最も顕著な特徴は、メイスのような尾の棍棒がないことである。ノドサウルス類は、代わりに骨皮で覆われた柔軟な尾を持ち、仙骨の盾を持っている。多くのノドサウルス科はまた、他の特徴の中でも、肩から外側に突き出たスパイクと、首に2つの装甲のある半輪を持っていた。3つ目の提案された科であるポラカンティダエ科は、ノドサウルス類に似ているが、より脆弱な装甲と異なる分布を持ついくつかの属を対象に設立された。しかし、近年の系統解析ではほとんど裏付けがなく、流動的である。[20] [42] [43]真正アンキロサウルス類は、ローラシア大陸に固有の真正アンキロサウルス類ではなく、ゴンドワナ大陸の白亜紀に生息していた少数の分類群からのみ知られるより小さなグループであるパランキロサウルス類を除く、アンキロサウルス類のすべてのメンバーを包含する。[44]

ミムーラペルタが最初に命名されたとき、カークランドとカーペンターは、この分類群をノドサウルス科の候補として位置付けたが、当時は比較対象となる分類群がわずかしかなかった。系統分類上の位置付けは1998年にヨーロッパのポラカンサス科に近いと考えられるようになったため変更されたが[5] 、この所属には疑問が持たれてきた。[7] [13] [8]マシュー・ヴィカリーオスら (2004) は、アンキロサウルス属をアンキロサウルス・インセルタエ・セディス (Ankylosauria incertae sedis ) と考えたが[45] 、トンプソンら(2011)による分岐論的解析では、ミムーラペルタはカークランドとカーペンターが当初理論づけた基盤的ノドサウルス科であることが示唆されている。[20] 2010年に未記載標品を用いた系統解析の結果、ミムーラペルタはポラカンサス科、ノドサウルス科、アンキロサウルス科の外側にある独自のグループにおいて、ガルゴイレオサウルスの姉妹タクソンとされた。 [7] 2018年にリベラ=シルバらが行った系統解析(以下に掲載)では、ミムーラペルタはサウロプリテスやドンヤンゴペルタとともにノドサウルス科の中で最も基底的なグループに属する基底的なノドサウルス類として同定されたが、ガルゴイレオサウルスはより派生した系統として同定され、両者の区別がさらに証明された。[43]あるいは、ミムーラペルタはノドサウルス科とアンキロサウルス科の両方に属さないアンキロサウルス類である可能性もある。[46]
リベラ・シルバら(左)とトーマス・レイヴンら(右)の系統解析により、ミムーラペルタはノドサウルス科またはアンキロサウルス科の基底的メンバーであることが判明した。[43] [47]
古生態学
モリソン層は浅海および沖積堆積物の層で、放射年代測定によると、基部で1億5630万年前(Ma )、上部で約1億5000万年前まで遡り、後期ジュラ紀のオックスフォード期後期、キンメリッジ期、およびティトニアン 期前期に位置付けられます。[48] [49]モリソン層の古環境は、雨期と乾期が明瞭で、平坦な氾濫原がある半乾燥環境だったと解釈されています。[6]植生は、川沿いの針葉樹、木生シダ、シダ(ギャラリーフォレスト)の森林から、アラウカリアに似た針葉樹ブラキフィラムなどの樹木が散在するシダサバンナまで多様です。[6]恐竜が生息していたモリソン盆地は、ニューメキシコ州からアルバータ州、サスカチュワン州にかけて広がり、ロッキー山脈のフロントレンジの前身が西へ隆起し始めた頃に形成されました。東向きの流域からの堆積物は、小川や河川によって運ばれ、湿地、湖、河道、氾濫原に堆積しました。

マイムーラペルタが最初に発見されたマイガット・ムーア採石場は、1億5189万年前から1億5247万年前のものと推定されており、モリソン層のブラッシー・ベイスン層に確実に位置づけられています。この地域の古環境は、豊かな植物相と高い地下水位を有する河畔生態系と解釈されており、採石場自体は季節的な水場における恐竜化石の摩耗堆積物と解釈されています。このため、この採石場はアメリカで最も化石の豊富な遺跡の一つとなっており、恐竜化石に加えて、植物、花粉、糞石、さらには皮膚の印象標本までもが保存されていることが特筆すべき点です。[50]採石場から産出される植物の化石には、スギナ、ソテツ、イチョウ目、チェカノフスキ目、そして葉、木、花粉の化石に代表される様々な針葉樹目植物が含まれます。[3]採石場から産出される無脊椎動物には腹足類や、最も顕著なものとして化石ザリガニが含まれます。[3]
脊椎動物については、Hulettia hawesiやMorrolepis schaefferiなど、驚くほど質の高い化石に基づいて 3 種類の魚類がこの採石場から記載されています。採石場から最も一般的な化石は恐竜の化石です。大型肉食獣脚類のアロサウルスと大型ディプロドクス科竜脚類のアパトサウルスは、この採石場で最も一般的な 2 種の恐竜であり、ミムーラペルタはサイトで 3 番目に多いです。[3]この採石場からは、獣脚類のケラトサウルス、竜脚類のカマラサウルスとディプロドクス、おそらくは小型鳥盤類のナノサウルスなど、他のいくつかの恐竜の化石も知られています。採石場内ではワニ類やカメ類などの他の脊椎動物の化石はほとんど見つからず、湖や池のような連続した水たまりが存在しなかったことを示しています。[50]
噛み跡が保存されているミムーラペルタと獣脚類アロサウルスの化石が、ステファニー・ドラムヘラーらによってマイガット・ムーア採石場から発見され、摂食の痕跡が残る他の骨とともに記述されている (2020)。[50]他の化石とは異なり、ミムーラペルタとアロサウルスの化石は条線を保存しており、これを測定して歯状突起(鋸歯) の幅を決定したところ、骨学的遺物から知られている採石場の2大捕食者であるアロサウルスとケラトサウルスのどの既知標本よりも大きな歯と体の大きさの外挿が得られた。この外挿は、アロサウルスの異常に大きい標本、またはトルヴォサウルスやサウロファガナクスのような別の大型分類群のいずれかと一致しているが、どちらも採石場内に化石材料を保存していない。この結果は、化石学的証拠のみに基づいてこの遺跡の既知の多様性を高めるか、あるいはアロサウルスが共食いを行っていたことを示す強力な証拠となるかのいずれかである。噛み跡のある様々な骨格要素の位置と栄養価に基づくと、ミムーラペルタは死後まもなく捕食されたか、あるいは死後まもなく腐肉食にされたと予測される。[50]
参照

{kind=link}
{kind=link}
.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
参考文献
- ^ abcdefghijklmnopqrst Kirkland, JI, & Carpenter, K. (1994). コロラド州西部の上部ジュラ紀モリソン層から発見された北米初の先白亜紀前期アンキロサウルス(恐竜類). Archived 2022-04-08 at the Wayback Machine Brigham Young University Geology Studies , 40 , 25-42.
- ^ Sullivan, Sharon (2007年9月24日). 「巨大な恐竜の骨が採石場で発掘される」www.aspentimes.com . 2022年8月18日時点のオリジナルよりアーカイブ。 2022年8月18日閲覧。
- ^ abcdefghi Foster, J., Hunt-Foster, R., Gorman, M., Trujillo, K., Suarez, C., McHugh, J., ... & Schoenstein, H. (2018). コロラド州西部モリソン層における大型恐竜骨床、マイガット・ムーア採石場の古生物学、化石化、堆積学—ジュラ紀後期恐竜の保存様式への示唆.インターマウンテン・ウェストの地質学, 5 , 23-93.
- ^ abc カーペンター, K.、マイルズ, C.、クロワード, K. (1998). 「ジュラ紀のアンキロサウルス(恐竜類)の頭蓋骨」Nature 393: 782–783.
- ^ abcdefghijklmnopqrs Hunt, AP, & Scheetz, RD (1998). Ankylosaur (Dinosauria) specimens from the Upper Jurassic Morrison Formation Archived 2022-08-10 at the Wayback Machine The Upper Jurassic Morrison Formation: An Interdisciplinary Study: Denver Museum of Natural History, Denver, USA: May 26–28, 1994 , 22 , 145-177.
- ^ abcd Foster, J. (2007). 『ジュラシック・ウェスト:モリソン層の恐竜とその世界』インディアナ大学出版局. pp. 327–329. ISBN 978-0-253-34870-8
- ^ abcde Kirkland, JI, Hunt-Foster, RK, Foster, JR, & Loewen, M. (2010). モリソン層の模式地から新たに発見されたジュラ紀後期の恐竜ミムーラペルタの骨格要素により、曲竜類の系統発生の再評価が可能になった。Journal of Vertebrate Paleontology , 30 , 116A
- ^ abcd Tremaine, K., D'Emic, M., Williams, S., Hunt-Foster, RK, Foster, J., and Mathews, J., (2015), Hanksville-Burpee Quarry 産の最新ジュラ紀(ティトニアン)モリソン層(Brushy Basin 層)産の Mymoorapelta maysi 角竜の新標本の古生態学的意味 [abs.]:Journal of Vertebrate Paleontology Program and Abstracts、p. 226。
- ^ abc Burns, ME, & Lucas, SG (2016). ニューメキシコ産アンキロサウルス(恐竜類:鳥盤類)骨皮類の生層序. Fossil Record 4: Bulletin 67 , 67 , 9.
- ^ Farlow, James O.; Coroian, Dan; Currie, Philip J.; Foster, John R.; Mallon, Jordan C.; Therrien, François (2022-07-11). 「景観におけるドラゴン:上部ジュラ紀モリソン層(米国)および上部白亜紀ダイナソーパーク層(カナダ)における大型肉食恐竜の生息数のモデル化」. The Anatomical Record . 306 (7): 1669– 1696. doi : 10.1002/ar.25024 . ISSN 1932-8486. PMID 35815600. S2CID 250422526.
- ^ ポール、グレゴリー・S. (2016). 『プリンストン・フィールド・ガイド・トゥ・ダイナソーズ』 プリンストン大学出版局. p. 252. ISBN 978-1-78684-190-2 OCLC 985402380
- ^ abcde Ősi, Attila; Prondvai, Edina; Mallon, Jordan; Bodor, Emese Réka (2016-07-20). 「アンキロサウルス類(恐竜類:鳥盤類)における摂食適応の進化における多様性と収束」 . Historical Biology . 29 (4): 539– 570. doi :10.1080/08912963.2016.1208194. ISSN 0891-2963. S2CID 55372674. 2022年10月7日にオリジナルからアーカイブ。2022年8月10日閲覧
- ^ ab Raven, TJ (2021).装甲恐竜(鳥盤類:甲羅亜綱)の分類学、系統学、生物地理学、大進化史(ブライトン大学博士論文).
- ^ Arbour, VM, & Currie, PJ (2015). アンキロサウルス類恐竜の尾棍は段階的に重要な特徴を獲得して進化した.解剖学ジャーナル, 227 (4), 514-523.
- ^ Brown, Caleb M.; Henderson, Donald M.; Vinther, Jakob; Fletcher, Ian; Sistiaga, Ainara; Herrera, Jorsua; Summons, Roger E. (2017-08-21). 「非常に保存状態の良い3次元装甲恐竜が、色彩と白亜紀の捕食者と被食者のダイナミクスに関する知見を明らかにする」Current Biology . 27 (16): 2514–2521.e3. Bibcode :2017CBio...27E2514B. doi :10.1016/j.cub.2017.06.071. hdl : 1721.1/118950 . ISSN 0960-9822. PMID 28781051. S2CID 5182644.
- ^ Hulke, JW (1887). VII. 1881年当時は不完全であったPolacanthus foxiiの背側の盾と内骨格の一部について記述した補足ノート。2022年4月9日にWayback Machineにアーカイブ 。Philosophical Transactions of the Royal Society of London.(B.)、(178)、169-172。
- ^ Burns, Michael E.; Currie, Philip J. (2014-06-07). 「アンキロサウルス(恐竜類、鳥盤類)の骨皮の外部構造と内部構造、そしてその系統的関連性」. Journal of Vertebrate Paleontology . 34 (4): 835– 851. Bibcode :2014JVPal..34..835B. doi :10.1080/02724634.2014.840309. ISSN 0272-4634. S2CID 85943454.
- ^ BLOws, WT (2001). 17. ポーラカンティン恐竜の皮膚装甲.装甲恐竜, 363.
- ^ abcd Arbour, Victoria M.; Burns, Michael E.; Currie, Philip J. (2011). 「A review of pelvic shield morphology in ankylosaurs (Dinosauria: Ornithischia)」 . Journal of Paleontology . 85 (2): 298– 302. Bibcode :2011JPal...85..298A. doi :10.1666/10-071.1. ISSN 0022-3360. S2CID 129637814. 2022年7月23日時点のオリジナルよりアーカイブ。 2022年8月10日閲覧。
- ^ abc Thompson, Richard S.; Parish, Jolyon C.; Maidment, Susannah CR; Barrett, Paul M. (2012-06-01). 「アンキロサウルス類恐竜(鳥盤類:甲羅亜目)の系統発生」. Journal of Systematic Palaeontology . 10 (2): 301– 312. Bibcode :2012JSPal..10..301T. doi :10.1080/14772019.2011.569091. ISSN 1477-2019. S2CID 86002282.
- ^ Tumanova, TA (1987). モンゴルの装甲恐竜. ソ連科学アカデミー.
- ^ Vickaryous, Matthew K.; Hall, Brian K. (2006). 「ナインバンドアルマジロ(Dasypus novemcinctus)の骨皮の形態と発達(哺乳類、異節上皮門、帯状疱疹亜綱) 」 . Journal of Morphology . 267 (11): 1273– 1283. Bibcode :2006JMorp.267.1273V. doi :10.1002/jmor.10475. PMID 17051548. S2CID 23338307. 2022年7月23日時点のオリジナルよりアーカイブ。 2022年8月10日閲覧。
- ^ Vickaryous, Matthew K.; Hall, Brian K. (2008). 「アリゲーター・ミシシッピエンシス(古竜類、ワニ亜科)の真皮骨格の発達と骨皮の相同性に関する考察」. Journal of Morphology . 269 (4): 398– 422. Bibcode :2008JMorp.269..398V. doi : 10.1002/jmor.10575 . PMID 17960802. S2CID 5927674.
- ^ Arbour, Victoria M.; Burns, Michael E.; Currie, Philip J. (2011). 「アンキロサウルス類(恐竜類:鳥盤類)の骨盤盾形態のレビュー」Journal of Paleontology . 85 (2): 298– 302. Bibcode :2011JPal...85..298A. doi :10.1666/10-071.1. ISSN 0022-3360. S2CID 129637814.
- ^ Carpenter, Kenneth (1984-12-01). 「北米白亜紀のサウロペルタ(アンキロサウルス類:ノドサウルス科)の骨格復元と生態復元」 . Canadian Journal of Earth Sciences . 21 (12): 1491– 1498. Bibcode :1984CaJES..21.1491C. doi :10.1139/e84-154. ISSN 0008-4077. 2020年9月2日時点のオリジナルよりアーカイブ。 2022年4月9日閲覧。
- ^ Prasad, Vandana; Strömberg, Caroline AE; Alimohammadian, Habib; Sahni, Ashok (2005-11-18). 「恐竜の糞石と草食動物および草食動物の初期進化」 . Science . 310 (5751): 1177– 1180. Bibcode :2005Sci...310.1177P. doi :10.1126/science.1118806. ISSN 0036-8075. PMID 16293759. S2CID 1816461. 2022年4月12日時点のオリジナルよりアーカイブ。 2022年4月9日閲覧。
- ^ カーペンター, ケネス (1997). 「アンキロサウルス類」. カリー, フィリップ J.、パディアン, ケビン (編). 『恐竜百科事典』. バークレー: カリフォルニア大学出版局. pp. 16–17.
- ^ ab Hill, Robert V.; D'Emic, Michael D.; Bever, GS; Norell, Mark A. (2015). 「白亜紀恐竜の複雑な鰓鰓器と鳥類の傍舌舌亜門の古代性:恐竜の鰓鰓器」.リンネ協会動物学誌. 175 (4): 892– 909. doi : 10.1111/zoj.12293 .
- ^ Vickaryous, Matthew K. (2006年12月11日). 「アルバータ州立恐竜公園で発見された白亜紀後期のノドサウルス科恐竜、エドモントニア・ルゴシデンス・ギルモアの頭蓋骨解剖に関する新情報」 . Journal of Vertebrate Paleontology . 26 (4): 1011– 1013. doi :10.1671/0272-4634(2006)26[1011:NIOTCA]2.0.CO;2. ISSN 0272-4634. S2CID 130696265. 2022年8月9日時点のオリジナルよりアーカイブ。 2022年8月10日閲覧。
- ^ Whitlock, J., Trujillo, K., & Hanik, G. (2018). モリソン層の恐竜の群集構造(アメリカ西部内陸部). Geology of the Intermountain West , 5 , 9-22.
- ^ Butler, RJ, & Barrett, PM (2008). 白亜紀の草食恐竜の分布に対する古環境の影響.自然科学, 95 (11), 1027-1032.
- ^ Coombs Jr, WP (1995). テキサス州下部白亜紀のノドサウルス類(恐竜類:鳥盤類). Journal of vertebrate Paleontology , 15 (2), 298-312.
- ^ Coombs, W. (1979). 「アンキロサウルス類(爬虫綱、鳥盤綱)の後肢の骨学と筋学」. Journal of Paleontology . 53 (3): 666–84. JSTOR 1304004.
- ^ Coombs, W. (1978). 「アンキロサウルス類(爬虫綱、鳥盤類)の前肢筋」. Journal of Paleontology . 52 (3): 642–57. JSTOR 1303969.
- ^ Coombs, Walter P. (1978-12-01). 「恐竜の行動適応に関する理論的側面」 . The Quarterly Review of Biology . 53 (4): 393– 418. doi :10.1086/410790. ISSN 0033-5770. S2CID 84505681. 2022年8月10日時点のオリジナルよりアーカイブ。 2022年8月10日閲覧。
- ^ Scheyer, Torsten M.; Sander, P. Martin (2004-12-10). 「アンキロサウルス類の骨皮の組織学:系統学と機能への示唆」Journal of Vertebrate Paleontology . 24 (4): 874– 893. doi :10.1671/0272-4634(2004)024[0874:hoaoif]2.0.co;2. ISSN 0272-4634. S2CID 86333501.
- ^ Arbour, Victoria M.; Currie, Philip J. (2015-08-31). 「アンキロサウルス類恐竜の尾棍は段階的に重要な特徴を獲得して進化した」. Journal of Anatomy . 227 (4): 514– 523. doi :10.1111/joa.12363. ISSN 0021-8782. PMC 4580109. PMID 26332595 .
- ^ Arbour, Victoria M.; Zanno, Lindsay E.; Evans, David C. (2022). 「アンキロサウルス科恐竜における種内戦闘の古病理学的証拠」. Biology Letters . 18 (12) 20220404. doi :10.1098/rsbl.2022.0404. ISSN 1744-957X . PMC 9727678. PMID 36475422.
- ^ Marsh, OC (1890). 新しい恐竜爬虫類の記載. American Journal of Science , 3 (229), 81-86. シカゴ
- ^ Marsh, Othniel Charles (1889-08-01). 「白亜紀の巨大角竜の発見」. American Journal of Science . s3-38 (224): 173– 176. Bibcode :1889AmJS...38..173M. doi :10.2475/ajs.s3-38.224.173. ISSN 0002-9599. S2CID 131187857. 2021年10月19日時点のオリジナルよりアーカイブ。 2022年8月10日閲覧。
- ^ Hone, DWE; Keesey, TM; Pisani, D.; Purvis, A. (2005-02-22). 「恐竜類における大進化の傾向:コープ則:恐竜はコープ則に従う」. Journal of Evolutionary Biology . 18 (3): 587– 595. doi : 10.1111/j.1420-9101.2004.00870.x . PMID 15842488. S2CID 14122343.
- ^ Zheng, Wenjie; Jin, Xingsheng; Azuma, Yoichi; Wang, Qiongying; Miyata, Kazuri; Xu, Xing (2018-02-27). 「中国のアルビアン-セノマニアン期に生息した最も基底的なアンキロサウルス亜科恐竜、尾棍の進化への影響」. Scientific Reports . 8 (1): 3711. Bibcode :2018NatSR...8.3711Z. doi :10.1038/s41598-018-21924-7. ISSN 2045-2322. PMC 5829254. PMID 29487376 .
- ^ abc リベラ=シルバ、エクトル E.;フレイ、エバーハルト。スタネスベック、ヴォルフガング。カーボット・チャノナ、ヘラルド。サンチェス・ウリベ、イヴァン・E.グスマン=グティエレス、ホセ・ルベン(2018)。 「メキシコ産白亜紀後期強竜類の古多様性とその系統学的重要性」。スイス古生物学ジャーナル。137 (1): 83–93。書誌コード:2018SwJP..137...83R。土井:10.1007/s13358-018-0153-1。ISSN 1664-2384。S2CID 134924657。
- ^ ソト=アクーニャ、セルジオ;バルガス、アレクサンダー O.カルーザ、ジョナタン。レッペ、マルセロ A.ボテーリョ、ジョアン F.パルマリベローナ、ホセ。サイモン・ガットシュタイン、カロライナ州。フェルナンデス、ロイ A.オルティス、ヘクトル。ミラ、ベロニカ。アラベナ、バルバラ。マンリケス、レスリー・ME。アラルコン・ムニョス、ジョナタン。ピノ、ファン・パブロ。トレヴィサン、クリスティーン(2021-12-09)。 「亜南極チリからの移行期のアンキロサウルスの奇妙な尾部兵器」。自然。600 (7888): 259–263。ビブコード:2021Natur.600..259S。doi :10.1038/s41586-021-04147-1. ISSN 0028-0836. PMID 34853468. S2CID 243794180. 2022年5月13日時点のオリジナルよりアーカイブ。 2022年8月10日閲覧。
- ^ Vickaryous, MK, Maryanska, T. ´ & Weishampel, DB (2004). Ankylosauria. DB Weishampel, P. Dodson & H. Osmolska (eds) ´The Dinosauria, 2nd edition. University of California Press, Berkeley. 363–392ページ
- ^ マジア、D.;アーバー、VM;カリフォルニア州ボイド。ファルケ、AA;クルザド・カバレロ、P.ワシントンDC、エヴァンス(2021年)。 「鳥盤類恐竜の系統命名法」。ピアJ。9 e12362。Bibcode :2021PeerJ...912362M。土井:10.7717/peerj.12362。PMC 8667728。PMID 34966571。
- ^ Raven, TJ; Barrett, PM; Joyce, CB; Maidment, SCR (2023). 「装甲恐竜(鳥盤類:甲羅亜綱)の系統関係と進化史」(PDF) . Journal of Systematic Palaeontology . 21 (1). 2205433. Bibcode :2023JSPal..2105433R. doi :10.1080/14772019.2023.2205433. S2CID 258802937.
- ^ Christiansen, EH; Kowallis, BJ; Dorais, MJ; Hart, GL; Mills, CN; Pickard, M.; Parks, E. (2015). 「モリソン層ブラッシー盆地層における火山活動の記録:北アメリカ西部の後期ジュラ紀への示唆」アメリカ地質学会特別論文513 : 1–XXX – ResearchGate経由。
- ^ Trujillo, Kelli; Kowallis, Bart (2015-01-20). 「米国西部内陸部、ジュラ紀後期モリソン層の再較正されたレガシー40Ar/39Ar年代」. Geology of the Intermountain West . 2 : 1–8 . doi : 10.31711/giw.v2.pp1-8 (2025年7月1日現在非アクティブ). ISSN 2380-7601.
{{cite journal}}:CS1 maint:DOIは2025年7月現在非アクティブです(リンク) - ^ abcd Drumheller, Stephanie K.; McHugh, Julia B.; Kane, Miriam; Riedel, Anja; D'Amore, Domenic C. (2020-05-27). 「高頻度の獣脚類の噛み跡は、ストレスのかかった後期ジュラ紀の生態系における摂食、腐肉食、そしておそらく共食いの証拠となる」. PLOS ONE . 15 (5) e0233115. Bibcode :2020PLoSO..1533115D. doi : 10.1371/journal.pone.0233115 . ISSN 1932-6203. PMC 7252595. PMID 32459808 .
