セイヨウニガナ
 |
| オリエンタルビタースイート | |
|---|---|
 | |
| 科学的分類 | |
| 王国: | 植物界 |
| クレード: | 維管束植物 |
| クレード: | 被子植物 |
| クレード: | 真正双子 |
| クレード: | ロシド類 |
| 注文: | ニシキギ目 |
| 家族: | ニシキギ科 |
| 属: | セイヨウニワトコ |
| 種: | C. orbiculatus |
| 二名法名 | |
| セイヨウニガナ | |
ニシキギ(Celastrus orbiculatus)はニシキギ科のつる植物です。 [ 1 ]一般にオリエンタルビタースイート、 [ 2 ] [ 3 ] [ 4 ]チャイニーズビタースイート、 [ 3 ]アジアンビタースイート、 [ 4 ]ラウンドリーフビタースイート、 [ 4 ]アジアティックビタースイートとも。
中国原産で、中国では最も広く分布しているニシキギ属の種であり、日本と韓国にも分布しています。[ 5 ]北米には1879年に導入され、[ 6 ]北米東部では侵略的外来種とみなされています。 [ 7 ]北米原産のニシキギ属の種であるニシキギ(Celastrus scandens)に酷似しており、容易に交雑します。[ 8 ]
説明
この落葉樹の特徴は、その蔓にあります。蔓は細く、ひょろ長く、銀色から赤褐色の樹皮を持ちます。蔓の直径は通常1~4cm(0.4~1.6インチ)ですが、成長を妨げなければ蔓は10cm(3.9インチ)を超えることもあり、切ると20年以上の年輪が現れることもあります。
セイヨウニガナは単独で生育すると茂みを形成し、樹木に近づくと、蔓が幹に巻き付き、高さ40フィートにも達します。巻き付いた蔓は、宿主となる樹木を絞め殺したり、過剰な重みで枝を折ったりすることが知られています。これは、成長の遅いアメリカ産のセイヨウニガナ(C. scandens )にも見られます。葉は丸く光沢があり、長さ2~12cm(0.8~4.7インチ)で、縁には鋸歯があり、蔓に沿って交互に生える。
腋生の集散花序に小さな緑色の花が咲く。果実は3つの弁を持つ蒴果で、裂開すると鮮やかな赤色の仮種皮が露わになり、種子を覆う。植物体全体が有毒である。[ 9 ]
- 完全なC. orbiculatus植物の例
- 植物の葉の形と根


{kind=link}
{kind=link}
{kind=link}
分布と生息地
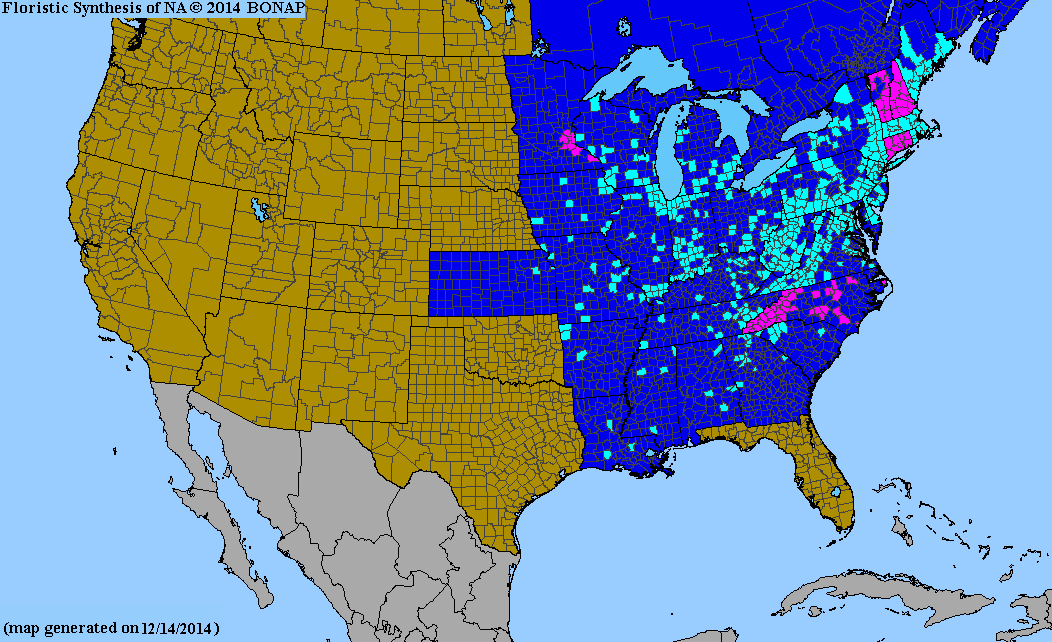
木材生産とレクリエーションを目的とした東部森林の組織的な撹乱により、オリエンタル・ビタースイートは北米東部の景観、道路脇、森林地帯に帰化しました。アメリカ合衆国では、南はルイジアナ州、北はメイン州、西はロッキー山脈に至るまで分布しています。[ 10 ] [ 11 ]湿潤林を好み、在来植物を覆い尽くすことが知られています。[ 12 ]
栽培
セイヨウニワトコは観賞用植物として栽培されており、英国では王立園芸協会のガーデン・メリット賞を受賞しています。[ 13 ]
侵略的外来種として
ニガナは生息環境において強力な競争相手であり、その拡散は他の多くの種の生存を脅かしてきました。この種の繁栄に貢献している特徴の一つは、魅力的な色の果実をつけることです。その結果、哺乳類や鳥類が食べ、種子を様々な場所に排泄します。
オリエンタルビタースイートが新しい地域に導入されると、在来植物にとって強力な競争相手が近隣に現れるため、地元の植物相が脅かされます。この種は東アジア原産ですが、米国には鑑賞目的で導入されました。[ 14 ]フラワーアレンジメントに使用され、不適切な廃棄のために無謀に地域に導入され、ジョージア州からウィスコンシン州までの33州以上とアパラチア山脈の一部の生態系に影響を与えています。[ 14 ]この生物は主に植物が繁茂した地域の周辺で成長するため、資源のフロンティアに容易にアクセスできます。オリエンタルビタースイートのさまざまな環境で生育する能力は、アパラチア山脈沿いの多くの植物種に有害であることが判明しており、時間の経過とともに西に向かって移動しています。[ 15 ] [ 16 ] [ 17 ]

セイヨウニガナは、多様な侵入・拡散戦略を用いて、外来地域において周囲の植物種との競争に打ち勝ちます。これが、本種の防除が困難である大きな理由です。[ 18 ]本植物の侵入は、生態学的、管理的、そして農業的に多様な問題を引き起こし、環境保全活動の焦点となっています。
非生物的要因への反応
日光は、ニガナにとって最も重要な資源の 1 つです。管理された実験によって実証されているように、ニガナは日光の量が多い環境でより速く成長します。28% を超える日光を受け取った集団の研究では、成長とバイオマスの量が増加しました。[ 19 ]この研究では、織物の層を使用して利用可能な日光の割合を制御しました。この実験では、ニガナがより多くの日光にさらされた場合、総生存長 (TLL、各植物の茎の生存長) が増加しました。[ 19 ]ニガナが 2% の日光にさらされると、TLL 比は低下しました。[ 19 ]ニガナは、2% ではなく 28% の日光にさらされると、バイオマスが 20% 増加する可能性があります。この植物の日光に対する強い反応は、より多くの日光を求めて争い、より多くの日光を受け取ることで他の種に勝つことができるため、侵入種としての役割と平行しています。オリエンタルビタースイートは、2%の日光にさらされると成長率が低下する(光合成能力の低下のため)が、それでも90%の生存率を示した。[ 20 ]実験データによると、オリエンタルビタースイートは、平均で0.8~6.4%の透過率の低光条件に耐える高い能力がある。[ 21 ]同属のアメリカンビタースイートと比較して、光の少ない生息地に置かれた場合、オリエンタルビタースイートは高さが増し、地上部バイオマスと葉の総量が増加することが判明した。[ 20 ] [ 21 ]オリエンタルビタースイートは、他の多くの競合種と比較して、日光を獲得する上でより優れた競合種である。
気温は、外来種であるニガヨモギの成長と発達に影響を与えるもう一つの変数です。他の外来種とは異なり、夏の高温は植物の成長を阻害することが示されています。ニガヨモギは、年間降水量の多い生息地で好ましい状態にあることも示されています。これは、メギやニシキギといった他の一般的な外来種とは対照的であり、年間降水量の多い環境に置かれると定着確率が低下することが示されているため、注目に値します。[ 22 ]
最近の研究で分析された他の侵入種と比較して、オリエンタル・ビタースイートは開発地域が大部分を占める景観でより多く見られました。[ 22 ]また、開けた生息地や放棄された生息地も、他の侵入種と比較して、本植物の拡散にプラスの影響を与えることが分かりました。[ 22 ]さらに、本種は環境の境界にある生息地で非常に好まれます。様々な環境条件で生息できるこの能力は、本植物の拡散に対する懸念を引き起こします。
生物的相互作用
相互的な相互作用
ニガナが在来植物種との競争に勝つ決定的な要因は、菌根菌、特にアーバスキュラー菌根菌と共生関係を形成する能力である。[ 23 ]ニガナの成長はリンの吸収に大きく依存している。最近の研究では、リン濃度の低い土壌にアーバスキュラー菌根菌が存在する場合、アーバスキュラー菌根菌が存在しない高リン濃度の土壌環境に植物を置いた場合よりも成長が大きいことがわかった。[ 23 ]この研究の結果は、ニガナが周囲から養分を効果的に吸収できるようにするための共生関係の重要性を示している。さらに、菌根との共生関係により、この侵入種は根のバイオマスのエネルギーをより少なく利用して必要な養分を吸収することができる。これは、オリエンタル・ビタースイートが効果的な侵略的種として機能する上で決定的な要因である可能性がある。なぜなら、オリエンタル・ビタースイートは、地下部バイオマスではなく地上部バイオマスに多くのエネルギーを割り当てることができるからである。この植物の侵略性に関する重要な点は、光合成能力と生殖能力に依存している。[ 23 ]菌類との共生関係はアーバスキュラー菌根菌とのみ形成され、外生菌根菌とはそのような関係は観察されていない。これらの研究は、適切な菌根が、植物がその環境で生存できるかどうかを決定づける強力な要因であることを示している。[ 23 ]また、研究は、「外来植物種は、自身の根だけでなく、近隣の植物の根の周囲の土壌中の微生物群集を変化させ、それによって植物種間の競争的相互作用を変化させることができる」という証拠も示している。[ 23 ]これは、ニガヨモギにとって重要な侵略的形質である可能性がある。なぜなら、この形質によって、ニガヨモギは地下の共生微生物の関係を変化させ、周囲の植物に悪影響を及ぼすからである。[ 23 ]しかし、この生物がこの形質を侵略的戦略として利用しているかどうかを判断するには、さらなる実験が必要である。
競争的な相互作用
オリエンタルビタースイートの侵略的な特性の一つは、エネルギーを効率的に利用して草丈を伸ばすことで、類似の植物に対して競争上の優位性を与えている。2006年に実施された研究では、同属のアメリカンビタースイートと比較して、オリエンタルビタースイートは草丈が高く、地上部バイオマスと葉の総量も増加していることが示された。[ 20 ]これは、オリエンタルビタースイートが全ての基準でアメリカンビタースイートを上回ったというわけではない。オリエンタルビタースイートと比較して、「アメリカンビタースイートは茎の直径、葉面積、葉量と茎の質量の比が大きく」、アメリカンビタースイートは茎の長さなどオリエンタルビタースイートが重視する植物特性よりも、植物の後方部分の成長に重点を置いていたことを示唆している。[ 20 ]これは、オリエンタルビタースイートが周囲の植物との競争に勝つ上で、草丈が大きな役割を果たしていることを考えると重要である。[ 20 ]茎の長さを中心に成長させることで、光を吸収する強い位置を確保できる一方で、日陰のような状態を作り出して周囲の植物に悪影響を及ぼします。
本種の蔓状形態は、周囲の植物に悪影響を及ぼすことも示されています。例えば、この形態学的特徴は、周囲の樹木を巻き付ける能力を助長し、樹木が氷害を受けやすくしたり、植物の重みで枝が損傷したりするなど、樹木に全体的に悪影響を及ぼすことが示唆されています。[ 24 ]さらに、研究では、オリエンタルビタースイートが周囲の植物から養分を吸い上げる能力があることが示唆されています。この研究では、この現象が様々な環境で発生することが明らかになっており、本植物の相対的な可塑性の向上と養分吸収量の増加の両方を示唆しています。[ 21 ]
ある研究では、ニガヨモギの存在が周囲の土壌のアルカリ度を上昇させることが観察された。これは多くの侵入植物種に見られる特徴である。[ 24 ]これにより必須栄養素の利用可能性が変化し、在来植物の栄養素吸収能力が阻害される。ニガヨモギと土壌のアルカリ度との関係は一貫しているが、この観察結果にはいくつかのメカニズムが提案されている。この植物の地上部バイオマスが大きいため、アンモニアよりも硝酸塩の優先的な吸収が要求され、土壌の硝化が促進される。また、ニガヨモギは高い陽イオン交換容量を持ち、これもまたより大きなバイオマスを支えている。これらの機能のいずれかがアルカリ度の上昇を説明できる可能性があるが、正確なメカニズムを特定するにはさらなる実験が必要である。[ 24 ]
交配
オリエンタルビタースイートがもたらすもう一つの大きな脅威は、アメリカビタースイートとの交雑です。アメリカビタースイートの雌とオリエンタルビタースイートの雄の間では容易に交雑が起こりますが、逆の交雑も稀に起こることが知られています。結果として生じた雑種は、繁殖能力が完全に備わっています。[ 25 ]理論的には、オリエンタルビタースイートの侵略がさらに悪化した場合、広範囲にわたる交雑によってアメリカビタースイートの個体群全体が遺伝的に混乱し、絶滅に至る可能性があります。[ 15 ]
管理
ニガヨモギが北米の生息地に侵入する影響を最小限に抑えるためには、その成長と拡散を厳密に管理する必要があります。保全活動を成功させるには、早期発見が不可欠です。これ以上の成長と拡散を抑えるために、地上部の植生を刈り取り、一般的な除草剤であるトリクロピルを葉に散布します。グリホサートも化学的に防除する方法です。これら2種類の除草剤は、通常、他の植物が標的とされるのを防ぐために晩秋に植物に直接散布されます。これらの手順は、毎年、または再生が見られるたびに繰り返す必要があります。[ 26 ]トリクロピルはほとんどの動物や昆虫種には無毒で、一部の魚種にはわずかに毒性がありますが、水中での半減期は1日未満であるため、野外で使用しても安全かつ効果的です。[ 26 ] [ 27 ] 機械的な方法も使用されていますが、根を完全に除去することが難しいため、それほど効果的ではありません。[ 28 ]また、この種の駆除に役立つ生物学的防除剤も存在しない。[ 29 ]機械的および化学的方法が使用されているが、それらは状況を一時的に改善するに過ぎない。
植物化学物質
ビセラファノールAは、ニガヨモギ(Celastrus orbiculatus)の樹皮から単離された神経保護作用のある二量体トリノルジテルペンである。[ 30 ]
用途
果実の毒性は弱いものの、一部の家畜は葉を食べて影響を受けない。丈夫で強靭な蔓は、かごの編み物に適した材料である。繊維質の樹皮は、丈夫な紐を作るのに用いられる。
参考文献
- ^ハウ D. (1955)。 「セラストラス属の改訂版」。ミズーリ植物園の年代記。42 (3)。ミズーリ植物園プレス: 215–302。Bibcode : 1955AnMBG..42..215H。土井:10.2307/2394657。JSTOR 2394657。
- ^ Lee, Sangtae; Chang, Kae Sun 編 (2015).韓国在来植物の英語名(PDF) . 抱川:韓国国立樹木園. p. 402. ISBN 978-89-97450-98-52019年3月15日閲覧–韓国山林庁経由。
- ^ a b Weeks, Sally S.; Weeks, Harmon P. (Jr.) (2011).インディアナ州と中西部の低木と木質つる植物:識別、野生生物としての価値、そして造園利用. ウェスト・ラファイエット:パーデュー大学出版局. p. 392. ISBN 9781557536105。
- ^ a b c Czarapata, Elizabeth J. (2005).中西部北部の侵略的植物:その識別と防除のための図解ガイド. マディソン:ウィスコンシン大学出版局. p. 45. ISBN 9780299210540。
- ^ Zhang, Zhixiang; Funston, Michele. 「Celastrus orbiculatus」 . Flora of China . 第11巻 – eFloras.org経由、ミズーリ植物園(ミズーリ州セントルイス)およびハーバード大学植物標本館(マサチューセッツ州ケンブリッジ)。
- ^ 「Celastrus orbiculatus – Oriental Bittersweet Vine」 . Conservation New England . 2016年3月18日時点のオリジナルよりアーカイブ。 2009年10月31日閲覧。
- ^ Kartesz, John T. (2014). 「Celastrus orbiculatus」 .北米植物アトラス(NAPA)による郡レベルの分布図. 北米生物相プログラム(BONAP).
- ^ White, Orland E.; Wray M. Bowden (1947). 「東洋とアメリカのビタースイートハイブリッド」. Journal of Heredity . 38 (4): 125– 128. doi : 10.1093/oxfordjournals.jhered.a105705 . PMID 20242535 .
- ^リチャード・H.ウヴァ、ジョセフ・C.ニール、ジョセフ・M.ディトマソ(1997年)『北東部の雑草』イサカ、ニューヨーク:コーネル大学出版局、 pp.336-337。
- ^ハッチソン、マックス (1990). 「植生管理ガイドライン:丸葉ビタースイート」 .植生管理マニュアル. イリノイ自然史調査. 2005年9月7日時点のオリジナルよりアーカイブ。
- ^ Kurtz, Cassandra M. (2018).米国北部森林におけるオリエンタルビタースイートの評価. ペンシルベニア州ニュータウンスクエア:米国農務省森林局北部研究ステーション. 2018年11月20日閲覧。
- ^ Archana Pande、Carol L. Williams、Christopher L. Lant、David J. Gibson. 「マップ代数を用いた侵入植物の中規模分布の特定:米国イリノイ州南部におけるCelastrus orbiculatusの事例」 (PDF) Biol Invasions:2006。 2010年6月12日時点のオリジナル(PDF)からアーカイブ。
- ^ 「Celastrus orbiculatus Hermaphrodite Group」 . rhs.org . 英国王立園芸協会. 2020年4月12日閲覧。
- ^ a b McNab, WH; Loftis, DL (2002). 「米国アパラチア山脈南部のオーク林におけるオリエンタルビタースイートの発生確率と生息地の特徴」 .森林生態学と管理. 155 ( 1–3 ): 45– 54. Bibcode : 2002ForEM.155...45M . doi : 10.1016/S0378-1127(01)00546-1 .
- ^ a b Jones, Chad C. (2012). 「侵入植物種の将来分布予測における課題」 .森林生態学と管理. 284 : 69– 77. Bibcode : 2012ForEM.284...69J . doi : 10.1016/j.foreco.2012.07.024 .
- ^ Albright, Thomas P.; Anderson, Dean P.; Keuler, Nicholas S.; Pearson, Scott M.; Turner, Monica G. (2009). 「導入の空間的遺産:米国南部アパラチア山脈におけるニシキヘビ(Celastrus orbiculatus)の分布」. Journal of Applied Ecology . 46 (6): 1229– 1238. Bibcode : 2009JApEc..46.1229A . doi : 10.1111/j.1365-2664.2009.01707.x .
- ^ a b NRCS . 「Celastrus orbiculatus」 . PLANTSデータベース.米国農務省(USDA).
- ^ Greenberg, CH; Smith, LM; Levey, DJ (2001). 「侵入性蔓植物の果実運命、種子発芽および生育 - 『待ち伏せ』戦略の実験的検証」 . Biological Invasions . 3 (4): 364– 372. Bibcode : 2001BiInv...3..363G . doi : 10.1023/A:1015857721486 . S2CID 6742817 .
- ^ a b c Ellsworth, JW; Harrington, RA; Fownes, JH (2004). 「日光下および日陰におけるCelastrus orbiculatus実生の生存、成長、ガス交換」. American Midland Naturalist . 151 (2): 233– 240. doi : 10.1674/0003-0031(2004)151[0233:SGAGEO]2.0.CO; 2 . JSTOR 3566741. S2CID 85822380 .
- ^ a b c d e Leicht SA, Silander JA (2006年7月). 「光質の変化に対する侵入種Celastrus orbiculatus(ニシキギ科)と在来種のC. scandensの異なる反応」Am. J. Bot . 93 (7): 972–7 . Bibcode : 2006AmJB...93..972L . doi : 10.3732/ajb.93.7.972 . PMID 21642161 .
- ^ a b c Leicht-Young, Stacey A.; Pavlovic, Noel B.; Grundel, Ralph; Frohnapple, Krystalynn J. (2007). 「形態学的特徴を用いた在来種(Celastrus Scandens L.)と侵略的種(C. Orbiculatus Thunb.)の識別」The Journal of the Torrey Botanical Society . 134 (4): 441– 50. Bibcode : 2007JTBS..134..441L . doi : 10.3159/07-RA-028.1 . JSTOR 20063940 . S2CID 86456782 .
- ^ a b cイバニェス, イネス; シランダー, ジョン A.; ウィルソン, アダム M.; ラフルール, ナンシー; 田中 信之; 津山 郁太郎 (2009). 「多変量予測による侵入植物種の潜在的分布」.生態学的応用. 19 (2): 359– 75. Bibcode : 2009EcoAp..19..359I . doi : 10.1890/07-2095.1 . PMID 19323195 .
- ^ a b c d e fレット、カーリー N.;デワルド、ローラ E.ホートン、ジョナサン (2011)。 「菌根と土壌リンがCelastrus orbiculatusの成長に影響を与える」。生物学的侵入。13 (10): 2339。Bibcode : 2011BiInv..13.2339L。土井: 10.1007/s10530-011-0046-3。S2CID 22309836。
- ^ a b c Leicht-Young, Stacey A.; O'Donnell, Hillary; Latimer, Andrew M.; Silander, John A. (2009). 「侵略的植物種Celastrus orbiculatusの土壌組成とプロセスへの影響」. The American Midland Naturalist . 161 (2): 219. Bibcode : 2009AMNat.161..219L . doi : 10.1674/0003-0031-161.2.219 . S2CID 12833005 .
- ^ Pavlovic, NB; Young, SL (2007). 「外来侵入性つる植物と在来種の同属植物の識別:形態、遺伝学、および交雑」(PDF) . 米国地質調査所、生態系健全性・修復部.
- ^ a b Pavlovic, N. Leicht-Young, S., Morford, D, & Mulcorney, N. (2011). 「オリエンタル・ビタースイートを燃やすべきか、燃やすべきでないか:火災管理者の難問」(PDF) . 米国地質調査所、ミシガン湖生態学研究ステーション、五大湖科学センター. 2013年3月9日時点のオリジナル(PDF)からアーカイブ。 2013年4月21日閲覧。
{{cite web}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ Ganapathy, Carissa (1997). 「トリクロピルの環境的運命」(PDF) . 農薬規制局 環境モニタリング・害虫管理部. 2009年6月13日時点のオリジナル(PDF)からアーカイブ。 2013年4月21日閲覧。
- ^ Swearingen, J., Reshetiloff, K., Slattery B., & Zwicker, S. (2002). 「大西洋中部自然地域への植物侵入者」 . 『大西洋中部自然地域への植物侵入者』. 国立公園局および米国魚類野生生物局. p. 82.
{{cite web}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ Dryer, Glenn D. (2003). 「Oriental Bittersweet: Element Stewardship Abstract」 . Wildland Weeds Management & Research Program, Weeds on the Web. 2008年10月23日時点のオリジナルよりアーカイブ。
- ^ Ly, Wang; J, Wu; Z, Yang; Xj, Wang; Y, Fu; Sz, Liu; Hm, Wang; Wl, Zhu; Hy, Zhang (2013年4月26日). 「(M)-および(P)-ビセラファノールA、ニセラツス由来の有望な神経保護活性を持つ二量体トリノルジテルペン」. Journal of Natural Products . 76 (4): 745–9 . Bibcode : 2013JNAtP..76..745W . doi : 10.1021/np3008182 . PMID 23421714 .
{kind=link}
外部リンク
- 種のプロフィール – セイヨウニガナ ( Celastrus orbiculatus ) 、米国国立農業図書館、国立侵入種情報センター。セイヨウニガナに関する一般情報とリソースを掲載しています。
