テニアテルム
| メデューサヘッド | |
|---|---|
 | |
| 科学的分類 | |
| 王国: | 植物界 |
| クレード: | 維管束植物 |
| クレード: | 被子植物 |
| クレード: | 単子葉植物 |
| クレード: | ツユクサ類 |
| 注文: | ポアレス |
| 家族: | イネ科 |
| 亜科: | ポイデア科 |
| 部族: | コムギ科 |
| 属: | テニアテルム・ネフスキー[ 1 ] |
| 種: | T. caput-medusae |
| 二名法名 | |
| テニアテルム・カプト・メデューサエ (L.)ネフスキー | |
| 同義語[ 2 ] | |
リスト
| |
{kind=link}
テニアテルム(Taeniatherum)は、ユーラシアおよび北アフリカに生息するイネ科植物の属である。 [ 3 ] [ 4 ] [ 5 ]
唯一認められている種はメデューサヘッド(Taeniatherum caput-medusae)で、南ヨーロッパと中央ヨーロッパ(ポルトガルからロシアのヨーロッパ)、北アフリカ(アルジェリア、モロッコ、チュニジア)、アジア(トルコ、サウジアラビアからパキスタン、カザフスタン)原産です。[ 2 ] [ 6 ] [ 7 ] [ 8 ] [ 9 ]また、オーストラリア南部、チリ、北アメリカの一部にも帰化しています。[ 10 ] [ 11 ] [ 12 ] [ 13 ]
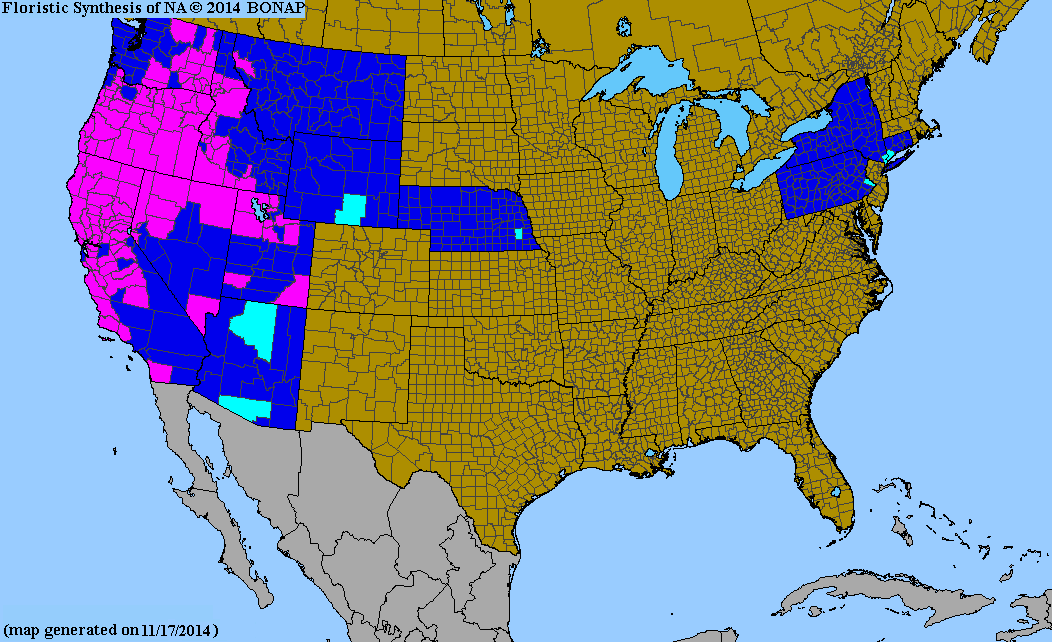
この侵略的な一年草である冬草は、北アメリカ西部の牧草地の生態系を変えつつあります。 [ 14 ]これは米国では1903年にオレゴン州でトーマス・ハウエルによって初めて観察されました。米国の陸地面積の48%は牧草地、牧草地、国立公園、自然保護区、その他の野生地域です。これらの土地は農業や生態系の完全性を守るために不可欠です。自然地域には、メデューサヘッドなど米国本土で自立した個体群として発生する多くの外来植物種が含まれています。2005年の時点で、メデューサヘッドは西部17州(南はノースダコタ州からテキサス州、西は太平洋岸まで)のおよそ972,700エーカー(3,936 km 2 )に蔓延しており、平均して年間12%の割合で広がっています。[ 15 ]メデューサヘッドが蔓延すると、過放牧された牧草地の在来植物を駆逐し、土地の価値を低下させ、山火事の危険を生み出す可能性があります。[ 16 ]
歴史と起源
メデューサヘッドは、カール・リンネによってElymus caput-medusaeとして初めて記載されました。ネフスキーは1934年にロシア産のメデューサヘッドを別の属Taeniatherumに分類することを推奨しました。1960年代には、カリフォルニア大学のジャック・メジャーが、地理的にも形態的にも異なる3つの分類群、T. caput-medusae、T. asperum、T. crinitumがあると示唆しました。ロシアを旅行した後、メジャーは北アメリカに導入された植物はTaeniatherum asperumに分類するのが適切だと考えました。この属は1986年にデンマークの科学者Signe Frederiksenによって改訂されました。彼は前述の異なる分類群をTaeniatherum caput-medusaeの亜種としました。[ 17 ]
亜種カプトメデューサエはヨーロッパ原産で、主にスペイン、ポルトガル、南フランス、アルジェリア、モロッコに分布しています。亜種クリニトゥムはギリシャとバルカン半島から東アジアにかけて分布し、亜種アスペルムは他の2亜種と完全に重複しています。[ 17 ]
アジアでは、メデューサヘッドはトルクメニスタン、イラン、シリア、そしてイスラエル北部に広く分布し、低山地や高原地帯に生息しています。農業用雑草であると同時に牧草地の雑草でもあります。窒素に富んだ土壌を好み、石や砂利の多い土壌でよく見られます。この雑草の炭化した種子は、イランの初期の農業遺跡から発見されています。種子は、羊や山羊の飼育が始まった初期の地層で初めて発見されました。[ 17 ]
ライフサイクルと生育習慣
メデューサヘッドは冬季一年草で、秋に発芽し、冬から早春にかけて根を張ります。根が早く伸びて土壌深部まで達するため、水分を巡って在来植物との競合を凌駕します。早春に開花し、6月か7月には小さな棘で覆われた種子が成熟します。棘は種子が家畜、人間、または通行する車両に付着するのに役立ちます。草が成長するにつれてシリカが蓄積されるため、ライフサイクルの初期を除いて家畜にとって口に合わないものになります。草は高密度の落葉層を形成しますが、シリカ含有量が多いため、落葉は他の植物よりもゆっくりと分解します。この落葉は在来植物の成長を抑制し、同時に自身の種子の発芽を促進します。そして数年後には大量の乾燥した燃料となり、山火事の原因となる可能性があります。[ 14 ]
メデューサヘッドの群落密度は、1平方フィートあたり数百から2,000本まで様々です。このばらつきは、年間降水量、土壌の種類、およびその地域の他の植生に直接関係しています。研究によると、メデューサヘッドは適応力が高く、1平方フィートあたり1,000本よりも1本の密度でより多くの種子を生産できることが示唆されています。[ 14 ]他のほとんどの一年草よりも成熟が遅いため、他の一年草が茶色のときに鮮やかな緑色をしていることが多いため、簡単に識別できます。成熟するにつれて、紫色の色合いに変わり、最終的には黄褐色になります。[ 14 ]この独特の植物季節学的特徴は、この侵入種の管理に利用できます。正規化植生指数(NDVI)値を使用して航空カラー赤外(CIR)画像における緑度を評価し、メデューサヘッドと他のより望ましい種を区別する最近開発された方法は、土地管理者が制御方法が必要な場所を決定するのに役立つ可能性があります。[ 18 ]
メデューサヘッドの種子は比較的短い距離しか散布せず、植物からの距離が離れるにつれて散布量は減少します。種子は移動物体への付着による散布に非常に適応しています。メデューサヘッドの種子散布期間が7月から10月と比較的長いのは、動物への付着の可能性を高めるための適応であると考えられます。[ 19 ]
識別
メデューサヘッドは高さ20~60センチメートルです。[ 20 ]細く弱い茎を持ち、基部で枝分かれすることがよくあります。小麦やライ麦に似た穂状の花序を持ちます。篩骨は長い芒を持ち、穎花は短い芒を持ち、種子の頭は層状の外観をしています。[ 21 ]芒は乾燥するとねじれ、あらゆる方向に広がり、神話に登場するメデューサの蛇に覆われた頭に似ています。芒の棘は種子が土壌に浸透するのを助けます。[ 22 ]穀物のような種子は土壌中で数年間生存することがあります。[ 14 ]
野生動物と放牧への影響
メデューサヘッドに侵食された土地の放牧能力は最大80%減少する可能性があります[ 23 ] 。野生生物の生息地と生物多様性も損なわれ、最終的には生態系機能の変化につながる可能性があります。メデューサヘッドがセージブラシに依存する種に与える影響についてはほとんど言及されていません。メデューサヘッドは、鳥類にとって重要な生息地を提供する植物群落を置き換えるため、キジライチョウ(Centrocercus属)の減少を悪化させる可能性があります[ 23 ] 。
ミュールジカやイワシャコなどの他の種は、クラゲ頭が良い食料源ではないため、クラゲ頭が繁茂している地域を避ける傾向があります。ミュールジカの場合、オレゴン州の研究では、クラゲ頭が広範囲に生息しているにもかかわらず、(他の植物群落と比較して)冬、夏、秋の餌となるミュールジカにとって、その地域は最も好まれず、春には順位が低かったことがわかりました。この摂食量の減少は、シカが食べないクラゲ頭の優勢と、それに伴う雑草の不足に関連していました。[ 24 ]イワシャコは、他に選択肢がない場合はクラゲ頭の種子(穎果)を摂取します。しかし、食べるクラゲ頭の種子をすべて自由に摂取できるようにすると、体重が大幅に減少しました。糞の中にはほとんど損傷を受けていない種子が見つかったため、鳥によるクラゲ頭の消化率は低かったことが示唆されました。[ 25 ]
制御方法
単一の防除方法ではクラゲを根絶することはできません。最良の結果を得るには、以下の2つ以上の方法を組み合わせた総合的病害虫管理( IPM)が必要となる場合が多くあります。
機械
プラウイングとディスク耕は、機械的な防除方法の2つです。どちらの方法もメデューサヘッドを効果的に防除し、翌生育期に発生を65%から95%削減できます。機械的な防除のみでメデューサヘッドを根絶することはほぼ不可能ですが、化学的な防除や植生回復と組み合わせることで、根絶の可能性は飛躍的に高まります。[ 14 ]
燃焼
放牧地の状態を改善する低コストの方法として、しばしば火入れが考えられています。クラゲ頭の防除において、火入れが注目されています。クラゲ頭が蔓延している地域の多くは、岩だらけであったり急勾配であったりして、他の対策が講じられないためです。カリフォルニア州では、火入れによるクラゲ頭の防除は様々な結果をもたらしました。地域によっては、火災後により望ましい植物群落が回復したのに対し、他の地域ではクラゲ頭が優勢な状態が続きました。クラゲ頭を焼却する際には、いくつかのガイドラインに従う必要があります。焼却は、晩春、種子が軟体状態(種子を握ると乳白色の物質が滲み出る状態)にあるときに行う必要があります。最初の火は、風に向かって燃やすことで容易に達成できる、ゆっくりと燃える火入れを行う必要があります。これにより、火が急速に燃え広がるのを防ぎ、その年の牧草が燃え尽き、クラゲ頭の穎果が死滅するのに十分な最高気温の期間を確保できます。生存可能なクラゲ頭の穎果は、ほぼ全て落葉中と土壌表面に存在します。[ 26 ]
焼却の有効性に関する過去の研究は、研究者に誤った期待を与えた可能性がある。穎果は温度依存的な後熟条件を厳しく持ち、成熟後約180日間は10℃以上の温度では発芽しない。これらの条件が満たされていれば、焼却区の落葉や土壌から採取された多くのクラゲ頭穎果は生存可能であった。しかし、後熟期間中に種子が発芽しなかったため、研究者は焼却によって実際よりも多くの成果が得られていると誤解していた。[ 26 ]
化学薬品
化学的な防除は、他の防除方法と併用することで効果を発揮します。グリホサート(ラウンドアップ)を種子ができる前の早春に0.375ポンド/エーカーで施用すると、良好な結果が得られます。このタイミングで施用することで、生育期の後半に成長する非標的種への被害も抑えられます。研究では、化学薬品を散布する前に火入れを行う方が、化学薬品のみを使用するよりも効果的であることが示唆されています。[ 14 ] 1960年代後半、米国のさまざまな地域でメデューサヘッド防除に対するパラコートの有効性を調べる研究が行われました。この研究では、パラコートはカリフォルニア州のメデューサヘッドの防除に効果的でしたが、ネバダ州リノでは草を抑制しないことがわかりました。これは、メデューサヘッドの蔓延に対して同時に散布して再播種するのに効果的な除草剤は存在しないことが判明したため、重要な意味を持ちました。[ 27 ]
生物学的
メデューサヘッドは、クラウンロット病や立枯れ病などの特定の根腐れ菌に感受性があることが判明しましたが、ベアパッチ病、褐色根腐れ病、および普通根腐れ病には感受性がありませんでした。これらの病気は根の総重量を減少させませんでしたが、立枯れ病は地上部のシュートの総乾燥重量を著しく減少させました。土壌伝染性病原体は、病気の環境条件が最適化されている限り、イネ科植物に深刻な影響を及ぼす可能性があります。立枯れ病は土壌水分の多い植物と関連しており、クラウンロット病と同様にメデューサヘッドに影響を与えました。対照的に、クラウンロット病は水ストレスを受けた植物に最も大きな影響を与えたため、米国西部の乾燥地域のイネ科雑草の効果的な生物学的防除となる可能性があります。また、ウエスタンウィートグラスなどの望ましいイネ科植物に重大な悪影響を与えなかったことも有望です。[ 28 ]
放牧
放牧のみでは、メデューサヘッドの駆除には適していません。最良の結果を得るには、放牧は統合プログラムの一部として行われます。放牧の時期と期間が適切に管理されている限り、放牧は効率的な管理手段となります。例えば、放牧を植生回復と併せて行う場合、放牧を行う前に望ましい草が定着していなければなりません。そうでなければ、植生回復は無駄になってしまいます。望ましい草が冬または早春までに成長期を終えている地域では、この時期に放牧を行うことでメデューサヘッドの駆除が可能です。まだ生育段階にあるため、家畜にとってより好ましい状態です。晩春、夏、秋の放牧は推奨されません。なぜなら、牛はメデューサヘッド以外の種を放牧するため、メデューサヘッドに競争上の優位性を与えてしまうからです。成熟したメデューサヘッド群落を防除するために家畜の放牧を行う場合、家畜を10日から2週間、飼育エリアに移動させ、雑草のないエリアに移動させる前に、雑草のない飼料を与える必要があります。これにより、家畜の体内を通過した種子が、メデューサヘッドのないエリアで発芽するのを防ぐことができます。[ 14 ]
復元
メデューサヘッドの管理計画には、必ず植生再生を組み込む必要があります。メデューサヘッドをまず駆除しなければ、蔓延した地域に再び種を蒔いても成功しません。既存のメデューサヘッドを駆除し、特に種子をこれ以上生産させないようにし、土壌中の種子バンクも縮小する必要があります。土壌水分や生育条件にもよりますが、通常2~3年かかります。その後、望ましい種の苗が定着します。耕起処理と除草剤散布を組み合わせることが、雑草を駆除し、望ましい植物の成長を促進する最も効果的な方法です。リステール、ブルーバンチウィートグラス、クレステッドウィートグラス、インターミディエイトウィートグラス、サーバーズニードルグラス、ニードルアンドスレッド、インディアンライスグラス、サンドバーグブルーグラス、シープフェスクはすべて、米国西部でメデューサヘッドに侵された地域を改修する際に効果的な競合的なイネ科植物です。[ 14 ]
防止
クラゲ頭の種子は、人間、動物、車両に付着して拡散することが多いため、蔓延地域においては、クラゲ頭の拡散を防ぐために、これらの交通を制限することが推奨されます。約3メートルの狭い封じ込め区域を設けることで、クラゲ頭の周辺地域への侵入を効果的に抑制することができます。[ 19 ]
大型多年生バンチグラスの密度が高い植物群落は、メデューサヘッドの侵入に対する抵抗力が高いため、大型多年生バンチグラスを促進・維持するための牧草地管理は、メデューサヘッドの蔓延を防ぐ上で非常に重要です。これにより新たな蔓延の発生は抑制されますが、効果的な管理には、新たな蔓延の発生源を探索し、根絶することも必要です。大規模な蔓延を抑制するよりも、新たな蔓延を抑制する方が効果的であり、多くの場合、より実現可能です。[ 23 ]
たとえ新たな蔓延を予防・抑制する試みが完全に成功しなくても、これらの努力は蔓延速度を遅らせ、研究者や土地管理者がより優れた予防、回復、抑制方法を開発するための時間を確保するのに役立ちます。また、蔓延速度を遅らせることは、蔓延リスクが最も高い地域の牧草地の健全性と生産性の向上にも役立ちます。積極的な予防プログラムがなければ、この雑草は蔓延を続け、生態系と経済への悪影響を増大させ続けるでしょう。メデューサヘッドの管理は1エーカーあたり高額に思えるかもしれませんが、蔓延を管理することで保護されるすべてのエーカーを考慮すると、その価格は非常に妥当です。さらに、土地価格の上昇を考慮すると、個々の畜産農家にとってメデューサヘッド管理にかかる費用は、メデューサヘッドの蔓延による生産損失を補うために追加の土地を購入する場合と比較して、急速に妥当なものになりつつあります。[ 23 ]
参考文献
- ^内: Trudy Sredne-Aziatskogo Gosudarstvennogo Universiteta。セリヤ 8b、ボタニカ17:38。1934 年。「名前 –テニアテルムネフスキー」。トロピコ。ミズーリ州セントルイス:ミズーリ植物園。2010 年4 月 15 日に取得。
タイプ標本: Taeniatherum crinitum (Schreb.) Nevski
- ^ a bキュー植物園指定植物科リスト
- ^ネフスキー、セルゲイ・アルセニェビッチ。 1934年。トゥルーディ・スレドネ・アジアツコゴ・ゴスダルストヴェンノゴ大学。セリヤ 8b、ボタニカ 17:38
- ^ Grassbase – 世界オンライン草本植物
- ^ Frederiksen, S. 1986. Taeniatherum (イネ科)の改訂Nordic Journal of Botany 6(4): 389–397
- ^アルテルヴィスタ フローラ イタリアーナ ジェネレテニアテラム
- ^ Danin, A. (2004). Flora Palaestina地域の植物分布地図帳: 1–517. イスラエル科学人文アカデミー, エルサレム
- ^ Cope, TA, Knees, SG & Miller, AG (2007). アラビア半島とソコトラ島の植物相 5(1): 1–387. エディンバラ大学出版局
- ^ Dobignard, D. & Chatelain, C. (2010).北アフリカの花の索引 1: 1–455。植物園および庭園植物園、ジュネーブ
- ^北アメリカの生物相 2013年郡分布図
- ^ 「Meghan Skaer、2014年1月24日、Global Invasions Network、『New Kid on the Block: A novel invader in an invaded community』」。2015年2月22日時点のオリジナルよりアーカイブ。 2015年2月22日閲覧。
- ^ Ausgrass2、オーストラリアの草、Taeniatherum caput-medusae
- ^ Zuloaga, FO、Morrone, O.、Belgrano, MJ、Marticorena, C. & Marchesi, E. (編) (2008)。南植物血管カタログ。ミズーリ植物園の系統植物学モノグラフ 107: 1–3348
- ^ a b c d e f g h i Zimmerman, JR他「メデューサヘッド:ネバダ州における経済的影響と制御」 ネバダ大学リノ校ファクトシートFS-02-37。
- ^ Duncan, CA, et al. (2004). 牧草地および野生地における侵入植物による経済的、環境的、社会的損失の評価. Weed Technology 18:1411–1416
- ^ドレーマン、クレイグ・C. フィールドノート No. 260、20ページ、カリフォルニア大学シエラ・フットヒル研究・普及センターの土壌サンプル、ブラウンズバレー、カリフォルニア州、2016年6月
- ^ a b c Kostivkovsky, V. and JA Young. (2000). 侵略的外来牧草地雑草:その原生生息地の一部を垣間見る. Rangelands 22:6 3–6.
- ^ Malmstrom, CM, HS Butterfield, L. Planck, CW Long, VT Eviner. (2017). 草地雑草被覆の動態と管理への反応を定量化する新たな微細航空マッピング手法. PLOS ONE 12(10): e0181665.
- ^ a b Davies, KW (2008). セイジブラシ草原植物群落におけるメデューサヘッドの分散と定着. Rangeland Ecology and Management 61: 110–115.
- ^ジェプソンマニュアルトリートメント
- ^ Stubbendieck, JL, et al. (2004).北米野生植物:フィールドガイド. ネブラスカ大学出版局.
- ^米国森林局の火災生態学
- ^ a b c d Davies, KW and DD Johnson. (2008). インターマウンテン西部におけるメデューサヘッドの管理は危機的状況にある. Rangelands . 30:13–15
- ^ Bodurtha, TS, et al. (1989). ミュールジカの生息地利用とバンチグラス群落における遷移の関係.野生生物管理ジャーナル53:2 314–319.
- ^ Savage, DE, et al. (1969). Chukar Partridges によるメデューサヘッドとダウニーブロムの穎果の利用. The Journal of Wildlife Management 33:4 975–978.
- ^ a b Young, JA, et al. (1972). 毎年繰り返される野焼きがクラゲ頭群集に及ぼす影響. Journal of Range Management 25:5 372–375.
- ^ Young, JA, et al. (1971). パラコートに対するメデューサヘッドの反応. Journal of Range Management 24:1 41–43.
- ^ Grey, WE, et al. (1995).根腐れ菌および冠腐れ菌を用いたブロムス( Bromus tectorum)およびメデューサヘッド( Taeniatherum caput-medusae )の生物学的防除の可能性. Weed Technology 9:2 362–365.
{kind=link}
外部リンク
- 種のプロフィール - メデューサヘッド(Taeniatherum caput-medusae)、国立侵略的外来種情報センター、米国国立農業図書館。メデューサヘッドに関する一般情報とリソースを一覧表示しています。
- 侵入草と覇権草ガイドのMedusahead (Taeniatherum caput-medusae )
- カリフォルニア大学バークレー校のCalPhotos写真データベースにあるTaeniatherum caput-medusae
