| リソウィシア 時間範囲:後期三畳紀
| |
|---|---|

| |
| 左上腕骨と大腿骨 | |
| 科学的分類 | |
| 王国: | 動物界 |
| 門: | 脊索動物 |
| クレード: | 単弓類 |
| クレード: | 獣弓類 |
| クレード: | †異常歯症 |
| クレード: | †ディキノドン類 |
| 家族: | †シュテッケリ科 |
| 亜科: | †プラセリナエ科 |
| 属: | †リソヴィシア ・スレジ & ニエドジヴィツキ、2019 |
| 種: | † L. ボジャニ
|
| 二名法名 | |
| †リソウィシア・ボジャニ スレイ&ニェジヴィエツキ、2019
| |
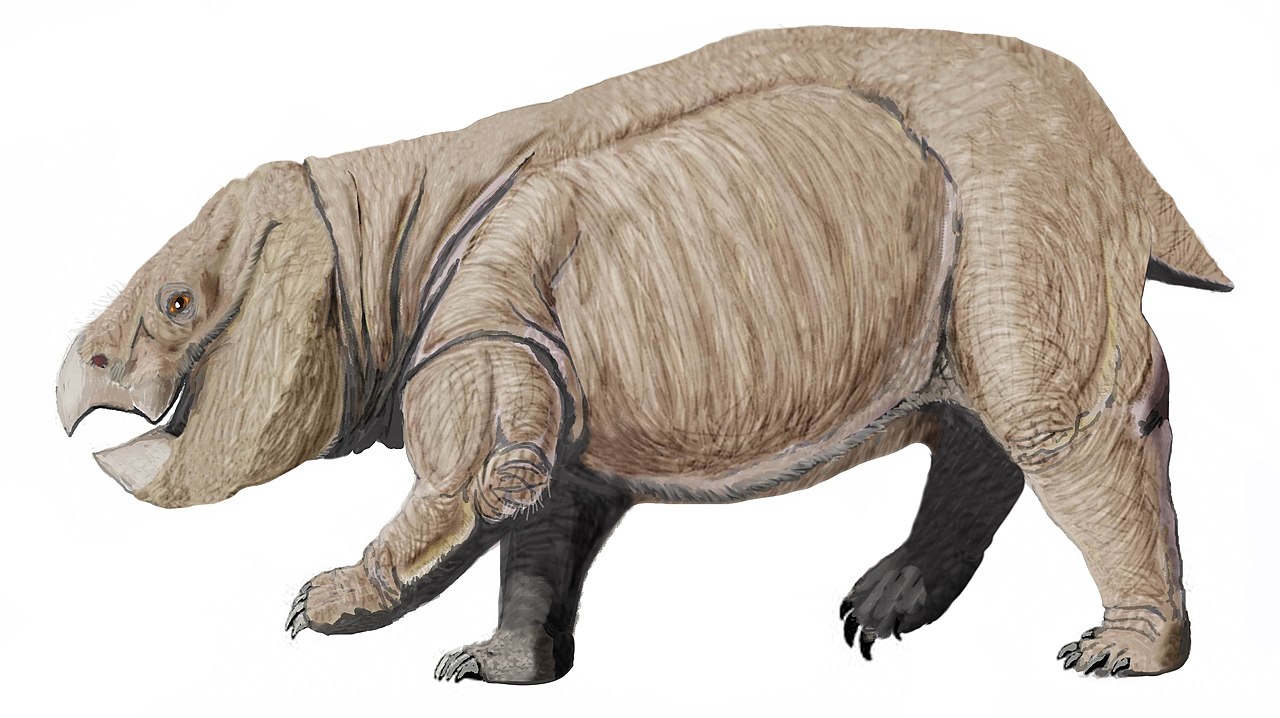
リソウィシアは、約2億1000万年から2億500万年前、後期三畳紀のノリアン後期またはレーティアン初期現在のポーランドに生息していた、絶滅した巨大な ディキノドン類の単弓類の属である。リソウィシアは既知のディキノドン類の中で最大であり、非哺乳類の単弓類としても最大で、全長約4.5メートル(15フィート)、直立すると腰高2.6メートル(8.5フィート)、体重は約5~7トン(5.5~7.7米トン)に達し、現代のゾウに匹敵する大きさである。また、三畳紀末の絶滅の直前まで生息していた最後のディキノドン類の一つでもあった。ポーランドでは2008年から巨大なディキノドン類の化石が発見されていたが、リソウィシアは2018年後半まで 新種として命名され、正式に記載されていなかった
リソウィキアは、四肢全てが体の真下に直立する直立姿勢で、ディキノドン類の中でも特異な存在です。これは現生哺乳類や恐竜の四肢に似ていますが、他の全てのディキノドン類(そして他の全ての非哺乳類単弓類)に典型的な、脚を伸ばした半直立姿勢とは異なり、大型哺乳類と独自に進化した多くの四肢の特徴を共有しています。リソウィキアはその生息環境において最大の動物であり、体の大きさや生態は、後期三畳紀のほとんどの生態系において大型の高地草食動物の役割を担っていた草食性の「前竜脚類」である竜脚形類に匹敵します。リソウィキアが発見される以前、ディキノドン類は竜脚形類との競争に敗れ、同様の大型体格を達成できなかったと考えられていました。リソウィシアは、ポーランド南部の生態系に竜脚形類がいなかったためにその大型化を進化させたのかもしれないし、あるいは竜脚形類の直接的な競争相手として行動したのかもしれない。
説明

ディキノドン類としては非常に大型であったものの、リソウィシアは北アメリカに生息する近縁種のプラセリアスなど、三畳紀の他のスタレッカー類とよく似ている。四足歩行で、丸々とした樽型の体、嘴状の顎を持つ大きな頭部、短い首、ずんぐりとした四肢、そして非常に短い尾を持っていた。[1]
頭骨は不完全で、様々な孤立した断片しか知られていないが、他のスタレッケリッド類と類似した頭骨を有していたと判断するには十分である。側頭窓は大きく、頭骨後部はその間の高い隆起に引き込まれ、頭骨後部に広範な顎筋の付着部を提供していた。吻は他のスタレッケリッド類と同様に比較的細長く、前方には歯のないカメのような嘴があった。他の三畳紀ディキノドン類と同様に、リソウィシアには完全に歯がなく、ほとんどのディキノドン類に典型的な牙さえもなかった。その代わりに、嘴の後ろの上顎骨から犬歯状突起と呼ばれる一対の短く太い三角形の突起があり、これは近縁のイスチグアラストアや他のスタレッケリッド類のものと類似していた。同様に、頭蓋骨後部の方形骨の比率をイスキグアラスティアのものと比較すると、リソウィシアの頭蓋骨は幅が広く、顎関節の各方形骨間の幅が約43センチメートル(17インチ)であったことが示唆される。[2]
背骨と肋骨はディキノドン類に典型的であるが、首の頚椎は側面に珍しい空洞が掘られていることで特徴的である。これは、気嚢のシステムと鳥類のような呼吸器系に関連する一部の恐竜の椎骨の穴に似ている。しかし、恐竜とは異なり、リソウィキアの椎骨(側腔)には気嚢のための開口部がなく、椎骨の内部は充実している。この特徴はディキノドン類の中でリソウィキアに特有であり、その機能は不明である。骨盤と後肢の骨は、動物の巨大な体格(例えば、知られている最大の大腿骨は80センチメートル(31インチ)の長さ)のため、他のディキノドン類と比較して非常に頑丈であるが、それ以外は他のスタレッカー類の骨と類似している。他のスタレッカー類と同様に、後ろ足も体の下に直立している。[1]
前肢
リソウィキアの前肢は、ディキノドン類の中でも非常に特徴的です。実際、その構造はディキノドン類だけでなく、非哺乳類 単弓類全体に特異であり、哺乳類や恐竜の四肢と共通の特徴を持っています。リソウィキアの前肢は体の下に完全に直立しており、肘関節は後方を向いているため、前腕は前後に振るだけで、傍矢状歩様をしています。この構造は、三畳紀の他の様々な大型カンネメイヤーリイフォーム類の後肢にも見られますが、それらはすべて、肘が外側に曲がったスプラウト状の前肢と、スプラウトした姿勢で歩くたびに橈骨と尺骨を回転させる関節を備えています。
この姿勢を実現するために、リソウィキアの前肢は、通常のディキノドン類と比較して、いくつかの顕著な解剖学的変化を経なければならなかった。上腕骨では、肩関節と肘が互いに平行であり、これは、寝そべるディキノドン類の上腕骨が回転するのとは異なり、肩の下、橈骨と尺骨の真上で上腕骨が直立している。前腕自体も、肘が体の真下、地面に近い位置にあるため、他のディキノドン類に比べて異常に短い。前肢の筋肉も、直立姿勢と歩行を容易にするために再配置され、四肢を前後に引く機能を持つようになり、寝そべる歩行のときのように上腕を回転させる能力は失った。[1]
サイズ

骨格は不完全だが、骨格の様々な要素が巨大であることから、リソウィキアは他のどのディキノドン類よりも大きな非常に大型の動物であったことがわかる。他のディキノドン類と骨を比較した結果、体長は約4.5メートル(15フィート)、股関節高は2.6メートル(8.5フィート)と推定された。[3] [4] [1]この上限推定値は、現存する最大の上腕骨部分に対応し、上腕骨全体の比率に基づくと、完全な状態であれば61.5センチメートル(24.2インチ)の長さになると推定される。[5]体重は当初、大腿骨と上腕骨の円周から約9.3メートルトン(10.3米トン)と算出されたが、これは四肢が支える必要があった重量の目安となる。しかし、リソウィキアではこれらの骨の軸は体の大きさに比べて不釣り合いに頑丈で太く、そのため体重は過大評価されていた可能性が高い。[1] [6]
その後、体重は体の全容積から質量をモデル化することで(体積モデル化)、より信頼性の高い推定が行われた。推定値は胸郭の周囲長と骨格周囲にモデル化された軟組織の量によって異なり、全体の平均重量は5.9トン(6.5ショートトン)、体脂肪やその他の組織が最小限の場合の最低推定重量は4.9トン(5.4ショートトン)、最大重量は最も重量が重い時で7トン(7.7ショートトン)とされた。この範囲は現代のアフリカゾウの重量に匹敵し、リソウィシアは哺乳類以外の単弓類としては最大であり、始新世に大型哺乳類(ブロントテア類など)が出現する前では最大の単弓類である。比較のために、大型の渦頭類であるタピノカニヌス は同じ方法を用いて平均重量892キログラム(1,967ポンド)と推定された。[6] [7]
発見の歴史

リソビエトの化石が、ポーランド南部シロンスク県ルブリニエツの町に近いリソビエトの村、リピエ・シロンスキエ粘土採掘場で発見された。リピエ・シロンスキエの化石を含む層は、主に緑色の泥岩でできている。リピエ・シロンスキエ粘土採掘場の年代は判定が難しく、生層序学的に異なる年代測定方法を用いると異なる結果が得られる。この遺跡は一般的にラエティアン期とされており、そうであればリソビエト恐竜は世界で最も若いディキノドン類の1つとなる。しかし、古生物学者のグジェゴシュ・ラッキとスペンサー・ルーカスは、リソビエト恐竜を含む大型脊椎動物の生層序に基づき、この遺跡はより古く、ノリアン中期期と推定されると主張している。彼らは、リソウィシアが他のディキノドン類よりも著しく若いわけではなく、世界の他の場所で最後に知られた属と同年代であると示唆した。[8]
リピエ・シロンスキエ粘土鉱山から採取された砕屑性ジルコンのサンプルを放射年代測定し、化石層の最大堆積年代を決定した。最も若いジルコン粒子は、後期ノリアン期に約2億1100万±300万年前の年代を記録したが、ジルコン結晶の形成時期と堆積物としての堆積時期の間に矛盾があるため、リピエ・シロンスキエの年代はこれよりも若い可能性があり、後期ノリアン期から前期ラエティアン期の推定を裏付けている。[9]
_Lisowicia_bojani.jpg/1280px-Szkic_wst%C4%99pnej_rekonstrukcji_(2008)_Lisowicia_bojani.jpg)
リソヴィキアの化石は、2006年に古生物学者のイェジー・ジク、トマシュ・スレイ、グジェゴシュ・ニェジヴィエツキによって初めて発見されました。彼らは当初、その巨大な体躯から竜脚形類恐竜の骨と誤認しました。その後、2008年に、部分的に関節した骨格と少なくとも2個体の複数の孤立した骨から、この化石が巨大なディキノドン類のものであると判明しました。2007年から2014年の間に、粘土採掘場から1,000点以上のリソヴィキアの骨が採取されました。 [10] [2] [11]
リソビシアは2019年1月まで正式に命名され、新分類群として公表されることはなかったが、その名称と記載は公式公表日より前の2018年11月にオンラインで公開された。属名は元の標本が発見された村リソビツェにちなんで名付けられ、種小名の bojaniは比較解剖学と古生物学の先駆的研究を行ったドイツの医師で博物学者ルートヴィヒ・ハインリヒ・ボヤヌス(1776-1827)に敬意を表して名付けられた。[10]リソビシアの既知の化石は、ポーランド科学アカデミー古生物学研究所とポーランドのワルシャワ大学古生物学・進化学科のコレクションに保管されている。
分類
リソウィシアはカンネメエリフォルム属で、三畳紀のディキノドン類のほとんどが属するグループであり、三畳紀後期に存在した唯一の現存するディキノドン類の系統であるスタレッカー科 に属している。特に、Sulej & Niedźwiedzki (2019) による2つの別々のデータセットを用いた系統解析では、どちらもリソウィシアがモロッコのモグレベリアと共に、北米でよく知られているプラセリアス属のプラセリアス亜科に近縁であることが判明した。同様の結果は、ポーランドのディキノドン類ウォズニケラの記載でSzczygielski & Sulej (2023) によって得られており、彼らはまた、南アフリカのペンタサウルスをプラセリア亜科の基底部で発見したが、ウォズニケラ自体はスタレッカー科の外側のより遠い位置で発見された。[12]
以下の系統樹は、シュチェジギエルスキ&スレイ(2023)の系統樹を修正・簡略化したもの。スタレッカー類とその近縁種との関係に焦点を当てており、最も短い(最も倹約的な)3つの系統樹の厳密なコンセンサスを示している。[12]




2024年、トマシュ・スレイはリソウィシアの骨学的記載において、カンネメイヤーリイフォルム類の非分岐論的系統樹を提唱した。多くの分岐論的分析のように形態のみに基づいて系統関係を判定するのではなく、スレイは地層学的および生物地理学的情報も用いて、時空間における祖先と子孫の関係を仮説化した。古生物学におけるこのアプローチは、ポーランドの古生物学者イェジー・ジクによって「クロノフィレティクス(年代系統学)」と名付けられた。[13]これらの結果は、上に示した系統樹のような従来の分岐論に基づく系統樹とは著しく異なる。スレイは、リソウィシアをローラシアのカンネメイヤーリイフォーム系統の最終個体と同定した。この系統は、中期三畳紀の「カンネメイヤーリイ科」シャーンベイカンネメイヤーリに始まり、ラビドサウルス、ウォズニケラ(分岐論的に復元されたより遠い位置とは対照的に)を経てプラセリアスを経て、リソウィシアに至った。上記の分岐樹とは異なり、彼はほとんどのスタレッカー亜科はプラセリア亜科と密接な共通祖先を共有しておらず、ローラシアのラディオドロムスとパラカンネメイヤーリイからゴンドワナ大陸に移住した中期三畳紀の「カンネメイヤーリイ科」の独自の系統から出現したと提唱した。[5]
スレイは、前頭骨と頭頂骨の形状、特に脳頭蓋を囲む正中線に沿った断面が、これらの系統を区別する重要な違いの一つであると主張した。リソウィシア属およびローラシア属の系統では、頭頂骨は垂直方向に細長く、三角形で、先端に向かって細くなっている。対照的に、シュテルレッカー亜属およびその系統では、頭頂骨は短く、下端はより幅広く丸みを帯びている。表面では、ローラシア属の系統では頭頂孔が頭蓋骨の後部により近く位置し、前頭骨はより長く、眼窩の縁に関与している。肩と肋骨が胸骨に単一に接合されているのも、この系統の特徴である。[5]
ディキノドン類の巨大化と進化

リソウィキアが発見される以前は、三畳紀の竜脚形類の巨大な体格への進化は恐竜特有の特徴であると考えられており、ディキノドン類を含む他の大型草食動物は同様のサイズには達していないと考えられていた。リソウィキアの象のようなサイズは、ディキノドン類でも巨大化が進化したことを示しており、後期三畳紀を通して進化するディキノドン類の体サイズの分析により、竜脚形類と同様に三畳紀のディキノドン類も体サイズが大きくなり、リソウィキアで頂点に達したことがわかった。実際、ディキノドン類はカーニアン期には一貫して竜脚形類よりも大きく、ノーリアン期からラエティアン期にかけてリソウィキアと同様に両者とも巨大なサイズを達成した。竜脚形類とリソウィキアの両方が同時期に巨大化を遂げたことは、これまで示唆されていた竜脚形類恐竜特有の特徴とは対照的に、生態学的要因が両系統の巨大化を可能にし、促進した可能性があることを示唆している。[1]
ディキノドン類の巨大化は、約2000万年かけて緩やかに進行し、筋骨格系が大きく再構築されて、リソウィキアの大きさと姿勢が実現した。リソウィキアの巨大化は、捕食者の体格の増大、もしくは、消化管の大型化によって食物からより多くの栄養分を得るための代謝適応によるものである可能性がある。 [14] [15]また、この研究は、後期三畳紀におけるディキノドン類の進化についてはまだ十分に解明されていない点も浮き彫りにしている。これまでは地理的に限定され、衰退していると考えられていたリソウィキアだが、この発見は、ディキノドン類が後期三畳紀においても依然として地理的に広く分布し、多様化を続け、特に巨大な体格を支える形質を進化させていたことを示している。[1]
古生物学
給餌と食事
リソウィキアは他の多くのディキノドン類と同様に草食動物であり、比較的長い嘴を使って咀嚼や噛みつきを行っていた。しかし、その大型の体躯は、このグループとしては典型的なものよりも高い位置の植物を摂食していたことを示唆している。一方、Sulej (2024) は、リソウィキアは低地の植物を摂食する「草食動物」であったと提唱した。この説は、眼窩の向きと後頭顆(頭骨と頸骨の接合部)の低い位置に基づいており、頭部を地面に向けて傾けた姿勢を常態としていたことを示唆していると解釈されている。この姿勢では、わずかに上向きの眼窩は、摂食時に地平線に向かって前方を向いていたと考えられる。[5]
リソウィシア属の糞石は、主に軟らかい植物や針葉樹を食べていたことを示しています。しかし、一部の糞石には木質物質が大量に含まれており、これは他のディキノドン類の糞石にも見られる特徴です。これは食性の季節変動を反映していると考えられており、おそらく好んで食べていた軟らかい植物が季節外れだった可能性があります。[14]
捕食

ディキノドン類リソウィシアや捕食性主竜 スモックが発見されたのと同じ場所で、幼いディキノドン類の骨に多数の噛み跡が見つかっており、幼いディキノドン類がスモックに優先的に、かつ積極的に狩られていたことを示している。ディキノドン類の骨片はスモックの糞石とされるものからも確認されており、リソウィシアのようなディキノドン類がスモックの食餌の主要成分であったことを示唆している。[15] [16]鼻骨が存在せず、他の吻骨が希少であることから、Sulej (2024) は、リソウィシアの吻は構造的に弱く、そのため捕食中、または吻を最初に死骸から分離させることによって、スモックのような捕食者に優先的に狙われたのではないかと示唆した。[5]
成長
リソウィキアの四肢骨の組織学的研究は、他の大型ディキノドン類と同様に、幼少期に急速な成長を示す特徴を示した。しかし、他のディキノドン類の成長速度は成長するにつれて遅くなるのに対し、研究されたリソウィキアの骨にはそのような成長減速の兆候は見られなかった。骨が推定よりも成体時の体格が大きい幼少期のものであった可能性は、すでに巨大な体格であったことに加え、骨が十分に骨化し発達的に成熟していたことから除外された。むしろ、リソウィキアは成体の大きさに達するまで中断することなく非常に急速に成長した可能性が高い。なぜなら、成長が遅く、低下した期間を示す明確な成長停止線(LAG)が見られなかったからである。LAGは本当に存在しなかったか、成体に向けての骨の大規模なリモデリングによって消去された可能性があり、どちらの場合も哺乳類や一部の恐竜と同様に、リソウィキアが恒久的に急速な成長を遂げていたことを裏付けるものである。[1] [10]
トイレ
リソウィキアのものとされる糞石は、かつて水たまりがあったと考えられる地域で、豊富に堆積して発見されています。これらの群集は、ブラジルで発見されたディキノドン類のディノドントサウルスが作ったとされる共同便所と類似しており、リソウィキアも同様の行動をとっていた可能性があります。共同便所は現代の群生性哺乳類にも記録されており、リソウィキアのようなディキノドン類が群れで生活していたという考えを裏付け、さらには現代の大型哺乳類のような複雑な社会行動をとっていたことを示唆しています。[14] [17]
古生態学

リソウィシアは、リピエ・シロンスキエにおいて、三畳紀後期の多様な動物と共存していた。これらの動物の大部分は主竜類であり、大型捕食性主竜類スモック、小型恐竜形類(シレサウルス類、コエロフィシスに似た新獣脚類) 、翼竜、小型ワニ形類(以前はポポサウルス上科と考えられていた)などが含まれていた。その他の小型爬虫類には、スフェノドント、分類不明の主竜形類、そしておそらくコリストデレなどが含まれる。リソウィシアと共存したことが知られている唯一の他の単弓類は、小型モルガヌコドント類の 哺乳類で、現生哺乳類に近縁であり、リソウィシアよりも桁違いに小型である。湿潤な淡水環境は、テムノスポンディル 両生類(カピトサウルス類と小型のプラギオサウルス類)や肺魚、シーラカンス、ヒボドンザメなどの豊富な魚類を支えていました。[8] [16]
リピエ・シロンスキエの環境は湿潤で沼地が多く、現代のエバーグレーズに匹敵する[14]。流れの緩やかな蛇行する川と三日月湖があり、豊かな植生に囲まれていた。その中には、ケイロレピディア科の針葉樹やイチョウ科、レピドプテリス属などの種子シダ、ソテツのアンドロストロバス属やビーニア属などが含まれていた。 対照的に、周辺の環境は乾燥して不毛であった可能性があり、これはおそらく乾生植物であるヒルメリエラの残骸や、火災の際に川によって運ばれた木炭の破片から示唆されている。[2] [18] [19]
大型の竜脚形類草食動物が存在しないのは不思議である。なぜなら、ドイツのプラテオサウルスのように、ヨーロッパの近隣地域を含む多くの後期三畳紀の生態系には、大型の竜脚形類草食動物が存在していたからである。リソウィキアのような巨大ディキノドン類が竜脚形類と直接競合していた可能性もあるが、彼らの不在は竜脚形類と無関係で、リソウィキアが巨大化して、他の生態系で竜脚形類が担っていた大型の草食動物としての生態学的役割を担った可能性もある。[3]
参考文献
- ^ abcdefgh Sulej, Tomasz; Niedźwiedzki, Grzegorz (2019年1月4日). 「直立四肢を持つ象サイズの三畳紀後期の単弓類」. Science . 363 (6422): 78– 80. Bibcode :2019Sci...363...78S. doi : 10.1126/science.aal4853 . ISSN 0036-8075. PMID 30467179.
- ^ abc Dzik、Jerzy;スレジ、トマシュ。ニエジヴィツキ、グジェゴシュ (2008)。 「ポーランドの最新三畳紀におけるディキノドン類と獣脚類の関連」。Acta Palaeontologica Polonica。53 (4): 733–738 .土井: 10.4202/app.2008.0415。
- ^ ab Dvorsky, George (2018年11月22日). 「三畳紀、このゾウサイズの哺乳類の近縁種は恐竜と並んで生きていた」Gizmodo . 2022年9月10日閲覧。
- ^ スタッフ (2018年11月23日). 「巨大哺乳類の『いとこ』発見」. Science Daily . 2022年8月22日時点のオリジナルよりアーカイブ。 2022年9月10日閲覧。
- ^ abcde Sulej、T. (2024)。 「三畳紀後期の巨大ジキノドン類リソウィキアの骨学と関係」。リンネ協会の動物学雑誌。202 (1)。 zlae085 。土井:10.1093/zoolinnean/zlae085。
- ^ ab Romano, Marco; Manucci, Fabio (2019年6月14日). 「Lisowicia bojaniのサイズ変更:後期三畳紀の巨大ディキノドン類の体積質量推定と3D復元」. Historical Biology . 33 (4): 474– 479. doi : 10.1080/08912963.2019.1631819 . ISSN 0891-2963. S2CID 196679837.
- ^ ロマーノ, マルコ; ルビッジ, ブルース (2019). 「タピノセファリッド恐竜類Tapinocaninus pamelae(単弓類:獣弓類)の初の3D再構成と体積体重推定」.歴史生物学. 33 (4): 498– 505. doi :10.1080/08912963.2019.1640219. ISSN 1029-2381. S2CID 203881268.
- ^ ab Racki, G.; Lucas, SG (2018). 「後期三畳紀のポーランドの特異な動物相とキュヴィエの絶滅論に基づくディキノドン類の絶滅時期」『歴史生物学』32 (4): 1– 11. doi :10.1080/08912963.2018.1499734. S2CID 91926999.
- ^ Kowal-Linka, M.; Krzemińska, E.; Czupyt, Z. (2019). 「上部三畳紀リピエ・シロンスキエ(リソヴィツェ)大陸堆積物(ポーランド)の最年少砕屑性ジルコン:リソヴィツェ骨含有層の最大堆積年代への示唆」.古地理学、古気候学、古生態学. 514 : 487– 501. Bibcode :2019PPP...514..487K. doi :10.1016/j.palaeo.2018.11.012. S2CID 134592379.
- ^ abc Gretchen Vogel (2018年11月22日). 「恐竜時代の巨人:古代の哺乳類のいとこはサイとカメの交配種のように見えた」 . Science . doi :10.1126/science.aaw1183. S2CID 135360687. 2019年12月22日閲覧。
- ^ St. Fleur, Nicholas (2019年1月4日). 「恐竜と並んで放牧していた象サイズの哺乳類の近縁種」ニューヨーク・タイムズ. 2019年1月6日閲覧。
- ^ ab Szczygielski、T.;スレジ、T. (2023)。 「Woznikella triradiata n. gen., n. sp. – 三畳紀後期のパンゲア北部および三畳紀のジキノドン類の世界的な分布に由来する新しいカンネメイエリフォーム・ディキノドン類」。コンテス・レンダス・パレボル。22 (16): 279–406 .土井: 10.5852/cr-palevol2023v22a16。
- ^ Dzik, J. (2005). 「年代系統学的アプローチ:不完全な化石記録に直面する成層化石学」(PDF) .古生物学特別論文集. 73 : 159–183 .
- ^ abcd Bajdek, P.; Owocki, K.; Niedźwiedzk, G (2014). 「ポーランドの上部三畳紀の推定ディキノドン類糞石」.古地理学・古気候学・古生態学. 411 : 1– 17. Bibcode :2014PPP...411....1B. doi :10.1016/j.palaeo.2014.06.013.
- ^ ab Niedźwiedzki, G.; Gorzelak, P.; Sulej, T. (2011). 「ディキノドン類の骨に残された咬痕と大型陸生捕食者の初期進化」Lethaia . 44 (1): 87– 92. Bibcode :2011Letha..44...87N. doi :10.1111/j.1502-3931.2010.00227.x.
- ^ ab Qvarnström, Martin; Ahlberg, Per E.; Niedźwiedzki, Grzegorz (2019). 「三畳紀の主竜類によるティラノサウルス類のような骨食作用」. Scientific Reports . 9 (1): 925. Bibcode :2019NatSR...9..925Q. doi :10.1038/s41598-018-37540-4. PMC 6353991. PMID 30700743 .
- ^ フィオレッリ、LE;エズクラ、メリーランド州。ヘッヘンライトナー、EM;アルガニャラス、E;タボルダ、JRA。ミネソタ州トロッティン。フォン・バツコ、MB;デソジョ、JB (2013)。 「既知の最古の共同トイレは、三畳紀の巨大草食動物の集団生活の証拠を提供している。」科学的報告書。3 (3348): 3348。ビブコード:2013NatSR...3.3348F。土井:10.1038/srep03348。PMC 3842779。PMID 24287957。
- ^ Jewuła, K.; Matysik, M.; Paszkowski, M.; Szulc, J. (2019). 「ポーランド南部、アッパーシレジアにおけるプラヤ、ギルガイ氾濫原、および河川環境の後期三畳紀の発達」.堆積地質学. 379 : 25– 45. Bibcode :2019SedG..379...25J. doi :10.1016/j.sedgeo.2018.11.005. ISSN 0037-0738. S2CID 133862052.
- ^ Wawrzyniak, Z. (2010). 「リピエ・シロンスク(ポーランド、上シレジア)産のディキノドン類(ディキノドン亜綱、獣弓類)の食性は? ― 後期三畳紀マクロフローラ研究の予備的成果」 Nowakowski, D. (編). 『化石脊椎動物の形態学と系統学』 DN Publisher, Wrocław. pp. 118– 124. OCLC 751467953.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
_Lisowicia_bojani.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
