

捕食とは、ある生物(捕食者)が別の生物(被食者)を殺して食べる生物学的相互作用である。これは、寄生や微小捕食(通常は宿主を殺さない)、そして寄生性(最終的には必ず宿主を殺してしまう)を含む一般的な摂食行動群の一つである。多くの捕食者も死骸を腐肉食するが、これは獲物の死骸を漁る(腐肉食)とは異なる。種子捕食者や破壊的な果食者も捕食者であるため、捕食は草食と重なる。
捕食行動は生物によって大きく異なります。多くの捕食者、特に肉食動物は、独自の狩猟戦略を進化させています。追跡捕食は獲物を積極的に探し出し、追跡する一方、待ち伏せ捕食者は獲物が捕獲の機会を待つため、しばしば隠密行動や攻撃的な擬態を用います。他の捕食者は日和見的または雑食性で、捕食行動は時折しか行いません。
ほとんどの絶対肉食動物は狩猟に特化しており、獲物を感知するために視覚、聴覚、嗅覚といった鋭い感覚を持っている場合があります。多くの捕食動物は、獲物を捕らえ、殺し、切り刻むために鋭い爪や顎を持っています。大型の肉食動物、例えば大型ネコ科動物は、より大きな獲物を仕留めるために通常、体力を必要とします。その他の適応としては、隠密行動、持久力、知性、社会行動、そして狩猟効率を高める攻撃的な擬態などが挙げられます。
捕食は被食者に強力な選択的影響を与え、被食者は警戒色、警戒音などのシグナル、カモフラージュ、防御力の高い種への擬態、防御用の棘や化学物質といった、捕食者に対する適応を発達させます。捕食者と被食者は、適応と反適応のサイクルである進化的軍拡競争に巻き込まれることがあります。捕食は少なくともカンブリア紀以降、進化の主要な原動力となってきました。
意味

最も基本的なレベルでは、捕食者は他の生物を殺して食べます。しかし、捕食の概念は広範で、文脈によって定義が異なり、多様な摂食方法が含まれます。さらに、獲物の死につながる関係性は必ずしも捕食とは呼ばれません。イチバチなどの寄生者は、宿主の体内または表面に卵を産みます。卵は孵化して幼虫となり、宿主を捕食し、宿主は必然的に死にます。動物学者は一般的にこれを寄生の一種と呼びますが、寄生者は従来、宿主を殺さないと考えられています。捕食者は生涯を通じて多くの獲物を捕獲するのに対し、寄生者の幼虫はたった1匹の獲物しか、あるいは少なくとも一度だけ餌が与えられるという点で、寄生者とは異なると定義できます。[1] [2]

他にも難しい、あるいは境界線上のケースがあります。微小捕食者は、捕食者と同様に他の生物だけを餌とする小動物です。生きた動物の血を吸うノミや蚊、生きた植物の樹液を吸うアブラムシなどがこれにあたります。しかし、通常宿主を殺さないことから、現在では寄生虫とみなされることも少なくありません。[3] [4]植物プランクトンや微生物の塊を食べる動物は、餌となる生物を消費して殺すため捕食者ですが、葉を食べる草食動物は、餌となる植物が通常攻撃を生き延びるため捕食者ではありません。[5]動物が種子(種子捕食または穀食)や卵(卵捕食)を食べる場合、生物全体を消費することになり、定義上、捕食者となります。[6] [7] [8]
腐肉食動物は、すでに死んでいる生物だけを食べる生物であり、捕食動物ではないが、ジャッカルやハイエナなど多くの捕食動物は、機会があれば腐肉食を行う。[9] [10] [5]無脊椎動物では、スズメバチなどの社会性スズメバチは、他の昆虫を狩ると同時に腐肉食動物でもある。[11]
分類範囲
哺乳類や鳥類における捕食者の例はよく知られていますが、[12]捕食者は節足動物を含む広範囲の分類群に見られます。昆虫ではカマキリ、トンボ、クサカゲロウ、サソリバエなどが一般的です。ハンノキバエなどの一部の種では、幼虫のみが捕食性です(成虫は食べません)。クモは捕食性であり、サソリ、ムカデ、一部のダニ、カタツムリ、ナメクジ、線虫、プラナリアなどの他の陸生無脊椎動物も捕食性です。[13]海洋環境では、ほとんどの刺胞動物(クラゲ、ヒドロ虫など)、有櫛動物(クシクラゲ)、棘皮動物(ヒトデ、ウニ、サンドドル、ナマコなど)、扁形動物が捕食性です。[14]甲殻類のうち、ロブスター、カニ、エビ、フジツボは捕食者であり、[15]甲殻類はほぼ全ての頭足動物(タコ、イカ、コウイカを含む)に捕食される。[16]

種子捕食は哺乳類、鳥類、昆虫に限られますが、ほぼすべての陸上生態系に見られます。[8] [6]卵捕食には、一部のコルブリッド ヘビなどの専門的な卵捕食者と、キツネやアナグマなどの卵を見つけると機会主義的に捕食する汎用動物の両方が含まれます。 [17] [18] [19]
ウツボカズラ、ハエトリグサ、モウセンゴケなどの一部の植物は肉食性で、昆虫を捕食します。[12]植物による捕食方法は多種多様ですが、多くの場合、餌となる罠、機械的刺激、電気刺激などを利用して最終的に獲物を捕獲し、捕食します。[20]一部の肉食菌は、締め付けリング状の能動的な罠、または粘着構造を持つ受動的な罠を使って線虫を捕獲します。[21]
多くの種の原生動物(真核生物)と細菌(原核生物)は他の微生物を捕食する。その摂食様式は明らかに古く、どちらのグループでも何度も進化してきた。[22] [12] [23]淡水と海洋の動物プランクトンでは、単細胞か多細胞かにかかわらず、植物プランクトンやより小さな動物プランクトンを捕食するのが一般的であり、多くの種のナノ鞭毛藻類、渦鞭毛藻類、繊毛藻類、ワムシ類、多様なメロプランクトンの動物の幼生、および2つのグループの甲殻類、すなわちコペポーダ類と枝角類で確認されている。[24]
採集

捕食者は餌を得るために、獲物を探し、追跡し、殺さなければなりません。これらの行動は採食サイクルを形成します。[26] [27]捕食者は獲物の地理的分布に基づいてどこで探すかを決定します。そして獲物を見つけたら、それを追跡するか、より良い選択肢を待つかを判断しなければなりません。追跡を選択した場合、その身体能力によって追跡のモード(例えば、待ち伏せや追跡)が決まります。[28] [29]獲物を捕らえた後、その処理(例えば、殺す、殻や棘を取り除く、そして飲み込む)にもエネルギーを費やす必要があるかもしれません。[25] [26]
検索
捕食者は、待ち伏せから活発なあるいは広範囲にわたる採餌まで、探索モードの選択肢を持っている。[30] [25] [31] [32]待ち伏せ法は、獲物が密集していて動きやすく、捕食者のエネルギー必要量が少ない場合に最適な方法である。[30]広範囲にわたる採餌はより多くのエネルギーを消費し、獲物が動かずまばらに分布している場合に用いられる。[28] [30]探索モードには連続体があり、移動間隔は数秒から数ヶ月に及ぶ。サメ、マンボウ、食虫鳥、トガリネズミはほとんど常に動いているが、造網クモ、水生無脊椎動物、カマキリ、チョウゲンボウはほとんど動かない。その中間では、チドリなどの海岸鳥類、クラッピーなどの淡水魚、テントウムシの幼虫(テントウムシ)は、積極的に探索と環境のスキャンを交互に行っている。[30]

獲物の分布はしばしば固まりになっており、捕食者は獲物が密集している場所を探し、その場所内を探索することで対応します。[ 25 ]餌がパッチ状になっている場合、例えばほとんど何もない海に魚の群れがほとんどいない場合など、捕食者は探索段階で各餌パッチを見つけるのにかなりの時間とエネルギーを費やして移動する必要があります。[33]例えば、マユグロアホウドリは定期的に約700キロメートル(430マイル)の範囲に餌を求めて飛行しますが、繁殖期には最大3,000キロメートル(1,860マイル)の範囲にまで餌を求めて飛行します。[a] [34]獲物が静止している場合、一部の捕食者は適切なパッチの位置を学習し、一定間隔でそこに戻って餌を食べることができます。[33]探索のための最適な餌探し戦略は、限界値定理を使用してモデル化されています。[35]
探索パターンはしばしばランダムに見える。その一例がレヴィ歩行で、短いステップの集合と時折長いステップを伴う傾向がある。これは、細菌、ミツバチ、サメ、そして狩猟採集民である人間など、様々な生物の行動によく当てはまる。 [36] [37]
評価

獲物を見つけた捕食者は、それを追いかけるか、それとも探索を続けるかを決めなければなりません。その決定は、それに伴うコストと利益によって決まります。昆虫を採餌する鳥は、探索に多くの時間を費やしますが、捕獲して食べるのは迅速かつ容易です。そのため、鳥にとって効率的な戦略は、見つけた美味しい昆虫をすべて食べることです。対照的に、ライオンやハヤブサなどの捕食者は獲物を見つけるのは簡単ですが、捕獲するには多大な労力を要します。この場合、捕食者はより選択的になります。[28]
考慮すべき要素の一つはサイズです。獲物が小さすぎると、エネルギーを大量に消費する割に合わない可能性があります。一方、大きすぎると捕獲が困難になる可能性があります。例えば、カマキリは前脚で獲物を捕らえますが、前脚は特定のサイズの獲物を捕らえるのに最適化されています。カマキリは、そのサイズから大きく離れた獲物を攻撃することに消極的です。捕食者の大きさと獲物の大きさには正の相関関係があります。[28]
捕食者は、ある場所を評価し、その中で獲物を探すのに時間を費やすかどうかを決定することがあります。[25]これには獲物の好みに関する知識が関係している可能性があります。例えば、テントウムシはアブラムシの獲物に適した植物の茂みを選ぶことができます。[38]
捕獲
捕食者は獲物を捕獲するために、公然と追跡する(追跡捕食)から近くの獲物に突然攻撃する(待ち伏せ捕食)まで、さまざまな追跡モードを持っています。[25] [39] [12]待ち伏せと追跡の中間にあるもう1つの戦略は弾道迎撃であり、捕食者は獲物の動きを観察し予測し、それに応じて攻撃を開始します。[40]
待ち伏せ
待ち伏せ型あるいは待ち伏せ型の捕食者は、ステルスや不意打ちで獲物を捕らえる肉食動物である。動物において、待ち伏せ型捕食は、捕食者が隠れた位置から獲物を見つけるまで周囲を偵察し、発見したら素早く狙いを定めた不意打ち攻撃を実行するという特徴がある。[41] [40]脊椎動物の待ち伏せ型捕食者には、カエルや、エンゼルシャーク、ノーザンパイク、イースタンフロッグフィッシュなどの魚類が含まれる。[40] [42] [43] [44]多くの無脊椎動物の待ち伏せ型捕食者には、陸上ではトタテグモやオーストラリアカニグモ、海ではシャコ類が含まれる。 [41] [45] [46]待ち伏せ型捕食者は、視野を狭める代わりに隠れるための巣穴を構築することが多く、隠蔽性を高める。一部の待ち伏せ型捕食者はまた、攻撃範囲内の獲物を引き付けるためにルアーを使用する。[40]攻撃は一度開始すると変更できないため、獲物を捕らえるには捕獲動作を迅速に行う必要がある。[40]
弾道迎撃
弾道迎撃とは、捕食者が獲物の動きを観察し、その動きを予測し、迎撃経路を算出し、その経路上で獲物を攻撃する戦略である。これは、捕食者が獲物の動きに合わせて攻撃を調整するという点で待ち伏せ捕食とは異なる。[40]弾道迎撃では、短時間の計画期間を要し、獲物に逃げる機会を与える。カエルの中には、ヘビが攻撃を開始するまでジャンプを待つものもいる。これにより、ヘビが攻撃を再調整する時間が短縮され、ヘビがリアルタイムでカエルを迎撃するために必要な角度調整が最大化される。[40]弾道捕食者には、トンボなどの昆虫、テッポウウオ(水噴射で攻撃)、カメレオン(舌で攻撃)、一部のナミヘビ科のヘビなどの脊椎動物が含まれる。[40]
追求
追跡捕食において、捕食者は逃げる獲物を追いかける。獲物が一直線に逃げる場合、捕食者が獲物より速いかどうかによってのみ捕獲が決まる。[40]獲物が逃げる際に方向転換する場合、捕食者は獲物に近づく際に平行移動などの新しい迎撃経路をリアルタイムで計算して追従する必要がある。 [40]多くの追跡捕食者は追跡を開始する前にカモフラージュを使って獲物にできるだけ気づかれずに近づく(ストーキング)。 [40]追跡捕食者には、人間、リカオン、ブチハイエナ、オオカミなどの陸生哺乳類、イルカ、シャチなどの海洋捕食者、マグロなどの多くの肉食魚類、[47] [48]ハヤブサなどの猛禽類(猛禽類)、トンボなどの昆虫などが含まれる。[49]
極端な追跡形態の一つに持久力狩猟、あるいは粘り強さ狩猟があります。これは、捕食者が獲物を長距離、時には何時間も追跡することで疲れさせる狩猟方法です。この方法は、人間の狩猟採集民や、アフリカリカオンやイエイヌなどのイヌ科動物によって用いられています。アフリカリカオンは極めて粘り強さが強い捕食者であり、比較的低速で何マイルも追跡することで獲物を疲れさせます。[50]
追跡捕食の特殊な形態として、ヒゲクジラの突進摂食があります。これらの非常に大型の海洋捕食者は、プランクトン、特にオキアミを捕食します。プランクトンの密集地帯に潜り込み、活発に泳ぎ回り、大量の水を飲み込み、羽毛状のヒゲ板を通して濾過します。[51] [52]
追跡型捕食者は、ライオンやオオカミのように集団で狩りをする社会的な動物もあれば、単独で狩りをする動物もある。 [2]
取り扱い
捕食者は獲物を捕らえると、それを慎重に扱わなければなりません。多くの捕食魚のように、鋭利な棘や毒のある棘を持つなど、食べるのが危険な獲物の場合は、非常に慎重に扱わなければなりません。イクタルリダ科などのナマズ類は、背中(背鰭)と腹(胸鰭)に棘を持ち、直立した状態で固定されます。捕獲されたナマズは暴れ回るため、これらの棘が捕食者の口に刺さり、致命傷を与える可能性があります。ミサゴのような魚食鳥類の中には、食べる前に獲物を引き裂くことで棘の危険を回避するものもあります。[53]
孤独な捕食と社会的捕食
社会的捕食では、捕食者の集団が協力して獲物を捕獲します。これにより、単独では倒せないほど大きな生物を捕獲することが可能になります。例えば、ハイエナとオオカミは協力してバッファローほどの大きさの草食動物を捕獲し、ライオンはゾウさえも狩ります。[54] [55] [56]また、獲物をフラッシュさせ、狭い範囲に追い込むなどの戦略によって、獲物をより容易に捕獲できるようにします。例えば、混合群れの鳥が餌を探しているとき、前方の鳥が昆虫を追い出し、それを後方の鳥が捕獲します。ハシナガイルカは魚の群れの周りに円を描いて内側へ移動し、魚を200倍に集めます。[57]チンパンジーは社会的に狩りをすることで、単独で狩れば容易に逃げるコロブスザルを捕獲することができ、協力するハリスホークはウサギを捕獲することができます。[54] [58]

異なる種の捕食者が協力して獲物を捕らえることもあります。サンゴ礁では、ハタやサンゴマスなどの魚類が、自分たちでは捕獲できない獲物を見つけると、オオウツボ、ナポレオンフィッシュ、タコなどに合図を送ります。これらの捕食者は小さな裂け目にも入り込み、獲物を捕獲することができます。[59] [60] シャチは、捕鯨船がヒゲクジラを捕獲するのを手伝うことが知られています。[61]
社会的な狩猟により、捕食者はより広範囲の獲物に取り組むことができるが、捕獲した食物をめぐる競争のリスクがある。単独行動の捕食者は、捕まえたものを食べる機会が増えるが、それを捕まえるためのエネルギー消費量が増え、獲物が逃げるリスクが高まる。[62] [63]待ち伏せ型の捕食者は、自身が獲物になるリスクを減らすために単独で行動することが多い。[64]食肉目(ネコ科、イヌ科、クマ科を含むグループ)の陸生種245種のうち、177種が単独行動であり、野生のネコ科動物37種のうち35種が単独行動であり、[65]ピューマとチーターもこれに含まれる。[62] [2]しかし、単独行動のピューマは他のピューマが獲物を分かち合うことを許容し、[66]コヨーテは単独行動と社会行動のどちらでもなり得る。[67]その他の孤独な捕食者には、ノーザンパイク[68] 、 オオカミグモ、節足動物の中の数千種の孤独なスズメバチ[69] [70]、そして多くの微生物や動物プランクトン[22]が含まれます。
専門分野
身体的適応
自然淘汰の圧力を受け、捕食者は獲物を検知し、捕獲し、殺し、消化するための様々な身体的適応を進化させてきた。これには、スピード、敏捷性、隠密行動、鋭い感覚、爪、歯、濾過器、そして適切な消化器系などが含まれる。[71]
捕食動物は獲物を探知するために、視覚、嗅覚、聴覚が発達している。[12]フクロウやハエトリグモなど多様な捕食動物は前向きの目を持ち、比較的狭い視野で正確な両眼視力を提供する。一方、獲物となる動物は全方位視力がそれほど鋭敏ではないことが多い。キツネなどの動物は、獲物が雪や土の下に2フィート(60cm)も隠れていても、その匂いを嗅ぎ分けることができる。多くの捕食動物は鋭敏な聴覚を持ち、反響定位を行う コウモリなど、音を能動的または受動的に利用して狩りをする動物もいる。[72]
大型ネコ科動物、猛禽類、アリなどの捕食動物は、強力な顎、鋭い歯、あるいは爪を持ち、獲物を捕らえて殺す。ヘビや、サギやウなどの魚食鳥類などの捕食動物の中には、獲物を丸呑みする者もいる。ヘビの中には、顎を自由に動かして大きな獲物を飲み込むことができるものもいる。また、魚食鳥類は槍のような長い嘴を持ち、素早く動き、滑りやすい獲物を突き刺して掴むことができる。[72]魚類などの捕食動物は、軟体動物の装甲殻を砕いたり、開いたりする能力を発達させている。[73]
多くの捕食動物は強靭な体格をしており、自分よりも大きな動物を捕らえて殺すことができます。これは、アリやトガリネズミなどの小型の捕食動物だけでなく、クーガーやライオンのような大きくて目に見えて筋肉質な肉食動物にも当てはまります。[72] [2] [74]
-
アカオノスリは鋭い鉤爪とくちばしを使って獲物を殺し、引き裂く。
-
専門家:魚を槍で突いたオオアオサギ
-
インドニシキヘビは顎を離して、このアクシスジカのような大きな獲物を飲み込む。






食事と行動
捕食動物は食性や狩猟行動において高度に特化していることが多く、例えば、ヨーロッパオオヤマネコは小型の有蹄類のみを狩ります。[75]ヒョウなどはより機会主義的なジェネラリストで、少なくとも100種を捕食します。[76] [77]スペシャリストは好みの獲物を捕獲することに高度に適応しているのに対し、ジェネラリストは好みの獲物が少ない場合に他の獲物に切り替える能力に優れている可能性があります。獲物が密集して(不均一に)分布している場合、獲物が目立ちやすく、より早く見つけられるため、捕食者にとって最適な戦略はより特化していると予測されます。[78]これは動かない獲物を捕食する捕食者には当てはまるようですが、動き回る獲物の場合は疑わしいです。[79]
サイズ選択的捕食において、捕食者は特定のサイズの獲物を選択します。[80]捕食者にとって、大きな獲物は厄介な存在となる可能性がありますが、小さな獲物は見つけにくく、いずれにせよ報酬も少なくなります。このことから、捕食者の大きさと獲物の大きさの間には相関関係が存在します。また、サイズは大きな獲物にとっての隠れ家となることもあります。例えば、成体のゾウはライオンによる捕食から比較的安全ですが、若いゾウは脆弱です。[81]
カモフラージュと擬態
ネコ科の動物、例えばユキヒョウ(樹木のない高地)、トラ(草原、葦の沼地)、オセロット(森林) 、スナドリネコ(水辺の茂み)、ライオン(開けた平原)などは、それぞれの生息地に適した色彩や特徴的な模様でカモフラージュされています。[82]
攻撃的な擬態において、昆虫や魚類を含む特定の捕食者は、色彩や行動を利用して獲物を引き寄せる。例えば、ホタル 科の雌は他の種の発光信号を模倣し、それによって雄のホタルを引き寄せて捕らえ、食べる。 [83] 花カマキリは待ち伏せ型の捕食者であり、ランなどの花に擬態して獲物をおびき寄せ、十分に近づいたら捕らえる。[84] カエルアンコウは非常によく擬態しており、頭の棒状の付属器の先端にエスカという餌を付けて、獲物を積極的に誘い寄せる。エスカを軽く振って小動物を模倣し、獲物が射程内に入ると非常に素早い動きで飲み込む。[85]
ヴェノム
ハコクラゲのような多くの小型捕食動物は、毒を使って獲物を捕らえます[86]。また、毒は消化を助ける働きもあります(ガラガラヘビや一部のクモの場合も同様です)。[87] [88]卵を捕食するように適応したマダラウミヘビは、毒腺が萎縮しており、3本指の毒の遺伝子に変異(2つのヌクレオチドの欠失)が生じて不活性化しています。これらの変化は、獲物を捕らえる必要がないという事実によって説明されます。[89]
電界

捕食魚類のいくつかのグループは、電場を感知して発生させることで獲物を検知し、追跡し、時にはヒキエイのように獲物を無力化する能力を持っています。[90] [91] [92]電気器官は改変された神経組織または筋肉組織に由来します。[93]
生理
捕食に対する生理学的適応には、捕食細菌が捕食する細菌の細胞壁から複雑なペプチドグリカンポリマーを消化する能力が含まれる。 [23]肉食脊椎動物は、5つの主要なクラス(魚類、両生類、爬虫類、鳥類、哺乳類)すべてにおいて、草食動物や雑食動物に比べて糖とアミノ酸の輸送速度が相対的に低い。これは、食事中の動物性タンパク質から多くのアミノ酸を摂取しているためと考えられる。 [94]
捕食者に対する適応
捕食に対抗するため、獲物は攻撃の各段階で使う防御手段を進化させてきた。[95] [12]獲物は、カモフラージュや擬態などを使って発見を回避しようとする。 [ 96]獲物は捕食者を発見し[98]他の捕食者にその存在を警告することができる。[99] [100] 発見された場合、攻撃の標的になることを避けようとする。例えば、有毒または不味いという合図を送る、[101] [102] [103]追跡しても利益にならないという合図を送る、[104] [105]あるいは群れを形成する。[106] [107]標的になった場合、装甲、針、不味さ、群がるなどの防御手段で攻撃をかわそうとする。 [108] [109] [110]そして、彼らは捕食者を驚かせたり、[111] [112] [113]死んだふりをしたり、尾などの体の一部を脱ぎ捨てたり、あるいは単に逃げたりすることで、攻撃から逃れることができることが多い。 [114] [115]
共進化

捕食者と獲物は天敵であり、彼らの適応の多くは互いに対抗するように設計されているように見える。例えば、コウモリは昆虫やその他の獲物を探知するための洗練されたエコーロケーションシステムを持ち、昆虫はエコーロケーションの鳴き声を聞き取る能力を含む様々な防御を発達させてきた。[116] [117]オオカミなど、陸上を走る多くの追跡捕食者は、獲物の速度増加に応じて長い手足を進化させた。[118]彼らの適応は進化的軍拡競争、つまり2つの種の共進化の例と特徴付けられる。[119]遺伝子を中心とした進化の見方では、捕食者と獲物の遺伝子は獲物の体をめぐって競合していると考えられる。 [119]しかし、ドーキンスとクレブスの「生命と食事」原理は、この軍拡競争は非対称であると予測しています。つまり、捕食者が獲物を捕まえることができなければ食事を失い、捕食に成功した場合は獲物が命を失うのです。[119]

軍備拡張競争という比喩は、攻撃と防御の絶え間ない進歩を暗示しています。しかし、こうした適応には代償が伴います。例えば、脚が長くなると骨折のリスクが高まります[120]。また、カメレオンの特殊な舌は、まるで弾丸のように水を舐める能力を持っていますが、水を舐めるには役に立たないため、カメレオンは植物の露を吸わなければなりません[121] 。
「命と食事」原理は、様々な理由から批判されてきた。自然選択における非対称性の程度は、適応形質の遺伝率に部分的に依存する。[121]また、捕食者が十分な量の食事を失うと、自らも命を失う。[120] [121]一方、失われた食事の適応度コストは予測不可能である。捕食者はより優れた獲物を素早く見つける可能性があるためである。さらに、ほとんどの捕食者は汎用性が高いため、特定の獲物の適応が捕食者に与える影響は小さい。特化は捕食者と被食者の共進化によって引き起こされるため、特化個体の希少性は、捕食者と被食者間の軍拡競争が稀であることを示唆している可能性がある。[121]
与えられた適応が、被捕食者の適応が捕食者の適応を促し、それが被捕食者のさらなる適応によって打ち消されるという共進化の結果であるのかどうかを判断することは困難である。別の説明としては、捕食者が競争相手、自身の捕食者、または危険な被捕食者に適応するエスカレーションが挙げられる。 [122]捕食への明らかな適応は、他の理由で生じた後、攻撃や防御に利用された可能性もある。コウモリの捕食対象となる昆虫の中には、コウモリが出現する前から聴覚が進化しており、縄張り防衛や交尾に用いられる信号を聞き取るために使用されていたものもある。[123]それらの聴覚はコウモリの捕食に応じて進化したが、コウモリにおける相互適応の明確な例はステルスエコーロケーションのみである。[124]
獲物が危険な場合、つまり捕食者に害を及ぼす棘や針、毒素、毒液を持つ場合、より対称的な軍拡競争が起こる可能性がある。捕食者は回避行動で対応することができ、それが擬態の進化を促す。回避行動は獲物との悪い経験から学習されることが多いため、必ずしも進化的な反応ではない。しかし、獲物が捕食者を殺す能力がある場合(サンゴヘビは毒でそうすることができる)、学習の機会はなく、回避行動は遺伝するしかない。捕食者は危険な獲物に対して対抗適応で対応することもある。北米西部では、一般的なガーターヘビがイモリの皮膚に含まれる毒素に対する耐性を獲得している。[121]
生態系における役割
捕食者は、自らの獲物を食べるという直接的な方法だけでなく、他の種による捕食を減らしたり、草食動物の採餌行動を変えたりするといった間接的な方法によっても生態系に影響を与えます。例えば、オオカミが川岸の植生に及ぼす生物多様性への影響や、ラッコがケルプ林に及ぼす生物多様性への影響などが挙げられます。これは、オオヤマネコやユキヒョウで観察される個体群動態のサイクルのような個体群動態への影響を説明できるかもしれません。[125] [126] [127]
栄養段階
捕食動物を分類する方法の一つは、栄養段階による分類である。草食動物を捕食する肉食動物は二次消費者であり、その捕食動物は三次消費者、というように分類される。[128]この食物連鎖の頂点には、ライオンなどの頂点捕食者がいる。[129]しかし、多くの捕食動物は食物連鎖の複数の段階から食物を食べており、肉食動物は二次消費者と三次消費者の両方を食べることがある。[130]これは、多くの捕食動物が、他の捕食動物に殺されて食べられるギルド内捕食と闘わなければならないことを意味する。例えば、コヨーテはハイイロギツネやボブキャットと競合し、時には彼らを殺してしまう。[131]
栄養段階の移転効率は、捕食によってエネルギーが上位の栄養段階にどれだけ効率的に伝達されるかを示す指標です。エネルギー伝達のたびに、捕食者が獲物を捕食する際に生じる熱、廃棄物、そして自然の代謝プロセスによって利用可能なエネルギーが減少します。その結果、ある栄養段階のエネルギーの約10%しか次の段階に伝達されません。これは、個々の生態系が支えることができる栄養段階の数を制限します。[132]
頂点捕食によって維持される生物多様性
捕食者は、単一の種が優占するのを阻止することで、生物群集の生物多様性を高める可能性があります。このような捕食者はキーストーン種として知られており、特定の生態系における生物のバランスに大きな影響を与える可能性があります。[133]この捕食者の導入または除去、あるいはその個体群密度の変化は、生態系内の他の多くの個体群の均衡に劇的な連鎖反応を引き起こす可能性があります。例えば、草原の草食動物は、単一の優占種が優占するのを阻止する可能性があります。[134]

イエローストーン国立公園からオオカミが排除されたことは、その栄養ピラミッドに深刻な影響を与えました。この地域では、オオカミはキーストーン種であると同時に頂点捕食者でもあります。捕食者がいなくなったことで、草食動物が多くの木本植物を過剰に食べ始め、その地域の植物群集に影響を与えました。さらに、オオカミはしばしば小川付近の動物の放牧を阻止し、ビーバーの食料源を守っていました。オオカミの排除はビーバーの個体数に直接的な影響を与え、彼らの生息地は放牧地となりました。捕食者の減少によりブラックテイル川沿いのヤナギや針葉樹の食害が増加し、減少したビーバーの個体数が水の流れを緩め、土壌を安定させることができなくなったため、水路の浸食が起こりました。このように、捕食者が生態系において極めて重要であることが示されました。[135]
人口動態
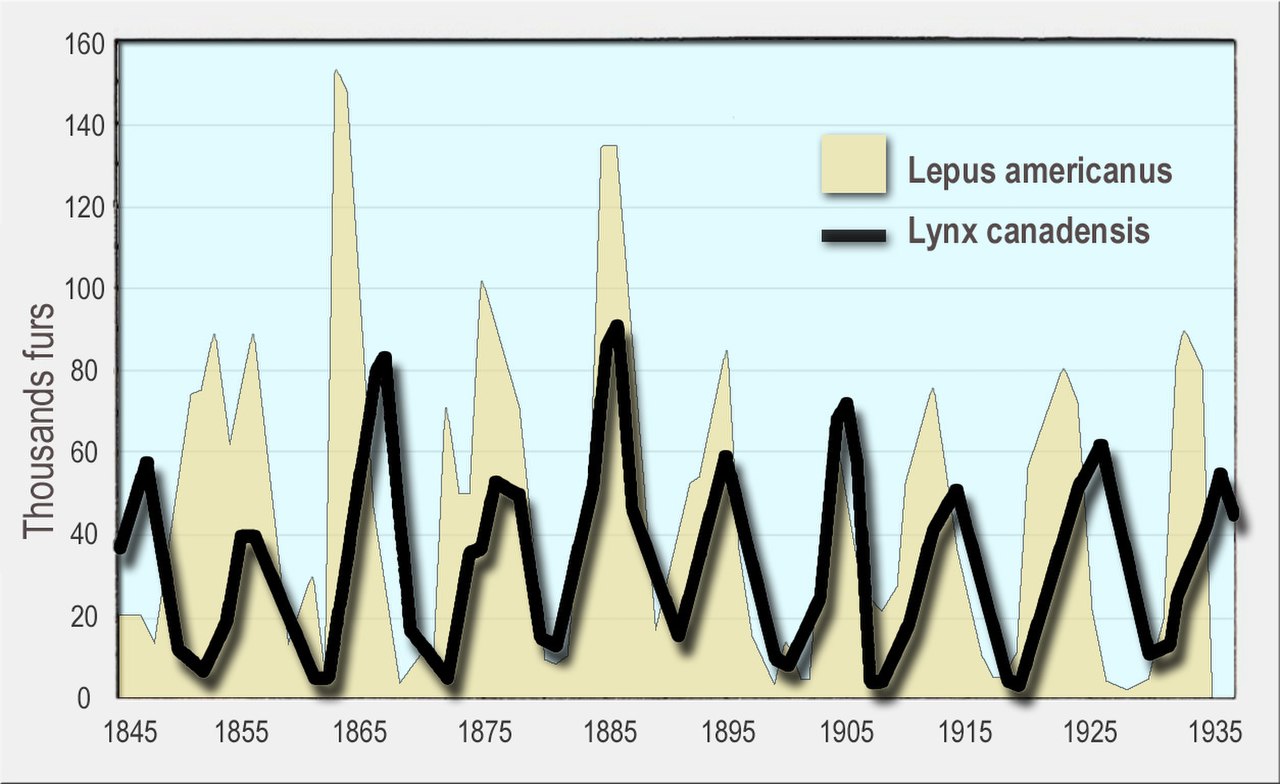
捕食者がいなければ、種の個体数は環境の収容力に近づくまで指数関数的に増加する可能性がある。 [136]捕食者は獲物を捕食することと、獲物の行動を変えることの両方によって、獲物の成長を制限する。[137]獲物の個体数の増減は、捕食者が産む子供の数の増加などを通じて、捕食者の数の増減にもつながる。
捕食者と被捕食者の個体群には周期的な変動が見られ、捕食者と被捕食者の周期の間にはしばしばずれが生じます。よく知られた例としては、ノウサギとオオヤマネコが挙げられます。アラスカとカナダの広大な北方林では、ノウサギの個体数はほぼ10年周期で変動し、オオヤマネコの個体数もそれに応じて変動します。これは、ハドソン湾会社の毛皮猟師が1世紀以上にわたって捕獲した動物の歴史的記録で初めて確認されました。[138] [127] [139] [140]
.png/1280px-Lotka-Volterra_model_(1.1,_0.4,_0.4,_0.1).png)
捕食者と被食者がそれぞれ1種ずつ存在するシステムの単純なモデルであるロトカ・ヴォルテラ方程式は、個体群循環を予測する。[141]しかし、このモデルの予測を実験室で再現する試みはしばしば失敗する。例えば、原生動物のディディニウム・ナストゥムをその被食者であるパラメシウム・カウダタムを含む培養物に加えると、パラメシウム・カウダタムはしばしば絶滅に追いやられる。[142]
ロトカ・ヴォルテラ方程式はいくつかの単純化仮定に基づいており、構造的に不安定である。つまり、方程式のいかなる変化も動態を安定化または不安定化させる可能性がある。[143] [144]例えば、一つの仮定は、捕食者が獲物に対して線形機能的反応を示すというものである。すなわち、捕食率は遭遇率に比例して増加する。この率が捕獲物の処理時間によって制限される場合、獲物の個体数は捕食者が制御できない密度に達する可能性がある。[142]もう一つの仮定は、すべての獲物の個体は同一であるというものである。実際には、捕食者は若く、弱く、病気の個体を選択する傾向があるため、獲物の個体数は再生可能である。[145]
捕食動物と被捕食動物の個体群を安定させる要因は数多くある。[146]一例として、複数の捕食動物、特に特定の被捕食動物が豊富な場合はその種に引き寄せられ、そうでない場合は他の場所を探す雑食動物の存在があげられる。[147]その結果、食物網がより単純な北部温帯および亜北極圏の生態系では個体群循環が見られる傾向がある。 [148]ノウサギとオオヤマネコのシステムは亜北極圏に生息するが、これにもコヨーテ、オオタカ、アメリカワシミミズクなどの他の捕食動物が関与しており、ノウサギが利用できる食物の変動によってサイクルが強化されている。[149]
ロトカ・ヴォルテラ・モデルの仮定を緩和した様々な数学モデルが開発されてきた。これらのモデルでは、動物が地理的に分布したり移動したりすること、性別や年齢構成など個体差があり一部の個体のみが繁殖すること、季節の変わり目など変化する環境に生息することなどが許容されている。[150] [151]また、2種以上の種の相互作用を同時に分析することも可能だ。このようなモデルは、捕食者と被食者の個体群動態が大きく異なり、しばしば混沌とすることを予測する。 [150] [152]被食者が捕食者から安全に避難できる避難場所の存在は、被食者がより大きな個体群を維持できるようにするかもしれないが、動態を不安定にすることもある。[153] [154] [155] [156]
進化の歴史
捕食は、一般に認識されている肉食動物の出現より数億年(おそらく数十億年)も前に遡ります。捕食は、様々な生物群で繰り返し進化してきました。[5] [157]約27億年前の真核細胞、約2億年前の多細胞生物、そして約6億年前~2億年前、おそらく約1億年前)の移動性捕食者の増加は、いずれも初期の捕食行動に起因するとされており、多くの非常に初期の化石には、小型捕食者種に起因すると思われる掘削穴やその他の痕跡の証拠が見られます。[5]これは、細胞、真核生物、有性生殖、多細胞性、サイズの増大、移動性(昆虫の飛翔を含む[158])、装甲殻や外骨格の出現など、主要な進化的変遷の引き金となったと考えられます。[5]
最古の捕食者は微生物であり、他の生物を包摂または捕食していました。化石記録が乏しいため、これらの最初の捕食者は1億年前から27億年前(10億年前)まで遡る可能性があります。[5]捕食はカンブリア紀の直前、つまり約5億5000万年前に顕著に重要になりました。これは、動物と藻類の石灰化がほぼ同時に発達したこと、 [159]と捕食を避けるための穴掘りが始まったことから明らかです。しかし、捕食者は少なくとも10億年前から微生物を捕食しており、[5] [160] [161]ほぼ同時期から選択的な(ランダムではない)捕食の証拠が見られます。[162]
オーロラルミナ・アッテンボローギは、イングランドのチャーンウッド・フォレストに生息するエディアカラ紀のクラウングループ刺胞動物(5億5700万年前~5億6200万年前、カンブリア爆発の約2000万年前)です。現代の刺胞動物と同様に、刺胞で小動物を捕食する、最古の捕食動物の一つと考えられています。 [163]
化石記録は、カンブリア紀以降、捕食者と獲物との長い相互作用の歴史を物語っています。例えば、一部の捕食者は二枚貝や腹足類の殻に穴を開け、他の捕食者は殻を破ってこれらの生物を捕食していました。[164] カンブリア紀の捕食者には、獲物を捕らえるのに適した付属肢、大きな複眼、そして昆虫の外骨格のような硬い素材でできた顎を持つアノマロカリス科のような無脊椎動物がいました。 [165]顎を持つ 最初の魚類には、シルル紀からデボン紀にかけて生息していた装甲を持ち、主に捕食性であった板皮類がいます。そのうちの一つ、体長6メートル(20フィート)のダンクルオステウスは、他の捕食者を捕食する世界初の脊椎動物「超捕食者」と考えられています。[166] [167]昆虫は石炭紀前期またはデボン紀後期に飛翔能力を発達させ、捕食者から逃れるなど様々な用途に利用されました。 [158] 史上最大の捕食動物には、白亜紀のティラノサウルスなどの獣脚類恐竜がいます。彼らはハドロサウルス類、角竜類、アンキロサウルス類などの草食恐竜を捕食していました。[168]
-
トンボ類の近縁種である石炭 紀の捕食性昆虫、メガネウラ・モニイは、陸生の捕食者から逃れるために飛ぶことができた。翼開長65cm(30インチ)というその大型は、当時、脊椎動物の空中捕食者が少なかったことを反映していると考えられる。
![オーロラルミナ・アッテンボロギは、エディアカラ紀の捕食者(約5億6000万年前)で、刺胞で獲物を捕らえる基底群の刺胞動物であった。[163]](https://upload.wikimedia.org/wikipedia/commons/thumb/6/63/Auroralumina_attenboroughii_reconstruction.jpg/1280px-Auroralumina_attenboroughii_reconstruction.jpg)


.png/1280px-Dunkleosteus_terrelli_(2024).png)


_with_its_prey.jpg){kind=link}
_with_its_prey.jpg/1280px-Polar_bear_(Ursus_maritimus)_with_its_prey.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.JPG){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
.jpg/1280px-Hawk_eating_prey_(cropped).jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_horiz.jpg){kind=link}
_horiz.jpg/1280px-Fig._1_-Riparian_willow_recovery_(26485120926)_horiz.jpg){kind=link}
{kind=link}
{kind=link}
.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
人間社会では
{kind=link}
{kind=link}

実用的な用途
雑食動物である人類はある程度捕食性であり、[169]武器や道具を使って魚釣りをし、[170]動物を 狩り、罠にかける。[171]また、イヌ、ウ、[172]ハヤブサなどの他の捕食種も利用して食用またはスポーツ用の獲物を捕らえる。[173]イヌとネコという 2種類の中型捕食動物は、西洋社会で最もよくペットとして飼われている動物である。[174] [175]南アフリカのサン族 などの人間の狩猟者は、持続狩猟を行う。これは追跡捕食の一種で、追跡者はクーズーなどの獲物よりも短距離では遅いかもしれないが、真昼の暑さの中で獲物が疲れ果てるまで追いかけ、最大5時間かかることもある。[176] [177]
生物学的害虫防除では、害虫の自然生息域から捕食者(および寄生者)を導入して個体群を制御しますが、予期せぬ問題を引き起こすリスクがあります。自然捕食者は、非害虫種に害を与えない限り、作物への被害を軽減する環境に優しく持続可能な方法であり、農薬などの化学薬剤の使用に代わる手段となります。[178]
象徴的な用途
{kind=link}
{kind=link}

映画では、プレデターが危険な人型の敵であるというアイデアは、1987年のSF ホラー アクション映画 『プレデター』とその続編3作で使用されている。[179] [180]恐ろしいプレデターである巨大な人食い ホオジロザメは、スティーブン・スピルバーグの1974年のスリラー映画『ジョーズ』でも中心的な存在となっている。[181]
捕食をテーマとした詩の中には、テッド・ヒューズの『パイク』のように、捕食者の意識が探求されているものがある。[182]アルフレッド・テニスンの1849年の詩『ああ、ああ、ああ』に登場する「歯と爪が赤い自然」というフレーズは、捕食者と獲物との闘争を指していると解釈されている。[183]
神話や民間の寓話では、キツネやオオカミなどの捕食動物の評判は複雑である。[184]キツネは古代ギリシャでは豊穣の象徴であったが、北欧では天候の悪魔、初期キリスト教では悪魔の生き物だった。イソップ物語以降では、キツネはずる賢く、貪欲で、ずる賢い存在として描かれている。[184]大きくて悪いオオカミは、 『赤ずきん』などの物語で子供たちに知られているが、アイスランドのエッダ・サガでは悪魔的な存在であり、終末論的な世界の終わりにオオカミのフェンリルが登場する。[184]中世には、男性がオオカミに変身する狼男の信仰が広まった。 [ 184]古代ローマや古代エジプトでは、オオカミは崇拝されており、ローマ建国の神話にはロムルスとレムスの乳を飲んでいる雌オオカミが登場する。[184]より最近では、ラドヤード・キプリングの1894年の『ジャングル・ブック』では、モーグリはオオカミの群れに育てられます。[184]北米のオオカミ、ハイイログマ、クーガーなどの大型捕食動物に対する態度は、積極的な迫害を伴う敵意や相反する感情から、20世紀後半には肯定的で保護的なものへと変化しました。[185]
参照
注記
- ^ 3000キロメートルの航続距離は、少なくとも往復6000キロメートルの飛行距離を意味します。
参考文献
- ^ Gurr, Geoff M.; Wratten, Stephen D.; Snyder, William E. (2012). 『生物多様性と害虫:持続可能な管理における重要課題』John Wiley & Sons. p. 105. ISBN 978-1-118-23185-2。
- ^ abcd Lafferty, KD; Kuris, AM (2002). 「栄養戦略、動物の多様性、そして体サイズ」. Trends Ecol. Evol . 17 (11): 507– 513. doi :10.1016/s0169-5347(02)02615-0.
- ^ Poulin, Robert ; Randhawa, Haseeb S. (2015年2月). 「収束線に沿った寄生の進化:生態学からゲノミクスへ」.寄生虫学. 142 (Suppl 1): S6 – S15 . doi :10.1017/S0031182013001674. PMC 4413784. PMID 24229807 .
- ^ プーリン、ロバート (2011). 「寄生虫への多くの道」. 『寄生虫学の進歩』第74巻. 第74巻. pp. 1– 40. doi :10.1016/B978-0-12-385897-9.00001-X. ISBN 978-0-12-385897-9. PMID 21295676。
- ^ abcdefg Bengtson, S. (2002). 「捕食の起源と初期進化」 Kowalewski, M.; Kelley, PH (編). 捕食の化石記録. 古生物学会論文集 8 (PDF) . 古生物学会. pp. 289– 317. 2008年9月10日時点のオリジナル(PDF)からアーカイブ。 2008年7月23日閲覧。
- ^ ab Janzen, DH (1971). 「動物による種子捕食」. Annual Review of Ecology and Systematics . 2 (1): 465– 492. Bibcode :1971AnRES...2..465J. doi :10.1146/annurev.es.02.110171.002341.
- ^ ニルソン、スヴェン G.ビョークマン、クリスター。フォースランド、パース;ジェイコブ・ヘグランド (1985)。 「島と本土の森林鳥群落における卵の捕食」。生態学。66 (4): 511–515。書誌コード:1985Oecol..66..511N。土井:10.1007/BF00379342。PMID 28310791。S2CID 2145031 。
- ^ ab Hulme, PE; Benkman, CW (2002). 「Granivory」. CM Herrera; O. Pellmyr (編). 『植物と動物の相互作用:進化論的アプローチ』. Blackwell. pp. 132– 154. ISBN 978-0-632-05267-7。
- ^ ケイン, アダム; ヒーリー, ケビン; ギレルム, トーマス; ラクストン, グレアム D.; ジャクソン, アンドリュー L. (2017). 「脊椎動物における腐肉食のレシピ ― 行動の自然史」.エコグラフィー. 40 (2): 324– 334. Bibcode :2017Ecogr..40..324K. doi :10.1111/ecog.02817. hdl : 10468/3213 . S2CID 56280901.
- ^ クルーク、ハンス (1972). 『ブチハイエナ:捕食と社会行動の研究』 カリフォルニア大学出版局. pp. 107– 108. ISBN 978-0-226-45508-2。
- ^ シュミット、ジャスティン・O. (2009). 「スズメバチ」.昆虫百科事典. pp. 1049– 1052. doi :10.1016/B978-0-12-374144-8.00275-7. ISBN 978-0-12-374144-8。
- ^ abcdef Stevens, Alison NP (2010). 「捕食、草食、そして寄生」. Nature Education Knowledge . 3 (10): 36.
- ^ 「捕食者、寄生虫、寄生虫類」オーストラリア博物館. 2018年9月19日閲覧。
- ^ 渡辺, ジェームズ・M. (2007). 「無脊椎動物の概要」. デニー, マーク・W.、ゲインズ, スティーブン・ディーン (編).潮だまりと岩礁の百科事典.カリフォルニア大学出版局. ISBN 978-0-520-25118-2。
- ^ Phelan, Jay (2009). What Is Life?: a guide to biology (Student ed.). WH Freeman & Co. p. 432. ISBN 978-1-4292-2318-8。
- ^ Villanueva, Roger; Perricone, Valentina; Fiorito, Graziano (2017年8月17日). 「捕食者としての頭足動物:行動の柔軟性、適応、そして摂食習慣の短い旅」. Frontiers in Physiology . 8 : 598. doi : 10.3389/fphys.2017.00598 . PMC 5563153. PMID 28861006 .
- ^ ハンセン、スヴェイン・アレ;エリクスタッド、ケル・アイナール (2012)。 「卵の捕食による長期的な影響」。行動生態学。24 (2): 564–569 .土井: 10.1093/beheco/ars198。
- ^ Pike, David A.; Clark, Rulon W.; Manica, Andrea; Tseng, Hui-Yun; Hsu, Jung-Ya; Huang, Wen-San (2016年2月26日). 「サーフ・アンド・ターフ:卵を食べるヘビによる捕食は、陸生トカゲにおける親の世話の進化につながった」. Scientific Reports . 6 (1) 22207. Bibcode :2016NatSR...622207P. doi :10.1038/srep22207. PMC 4768160. PMID 26915464 .
- ^ Ainsworth, Gillian B.; Calladine, John; Martay, Blaise; Park, Kirsty; Redpath, Steve; Wernham, Chris; Wilson, Mark; Young, Juliette (2016). 「捕食を理解する:スコットランドにおける近年の野鳥個体群変動とその要因に関する自然科学と地域知識を結集したレビュー」スコットランド・ムーアランド・フォーラム. doi :10.13140/RG.2.1.1014.6960.
- ^ ヘドリッヒ・ライナー;福島健二(2021年5月20日)「肉食性の起源について:動物食植物の分子生理学と進化」Annual Review of Plant Biology 72 (1). annurev–arplant–080620-010429. Bibcode : 2021AnRPB..72..133H. doi : 10.1146/annurev-arplant-080620-010429 . ISSN 1543-5008. PMID 33434053. S2CID 231595236.
- ^ Pramer, D. (1964). 「線虫捕獲菌類」. Science . 144 (3617): 382– 388. Bibcode :1964Sci...144..382P. doi :10.1126/science.144.3617.382. JSTOR 1713426. PMID 14169325.
- ^ ab Velicer, Gregory J.; Mendes-Soares, Helena (2007). 「細菌捕食者」. Cell . 19 (2): R55 – R56 . doi : 10.1016/j.cub.2008.10.043 . PMID 19174136. S2CID 5432036.
- ^ ab Jurkevitch, Edouard; Davidov, Yaacov (2006). 「捕食性原核生物の系統学的多様性と進化」.捕食性原核生物. Springer. pp. 11–56. doi :10.1007/7171_052. ISBN 978-3-540-38577-6。
- ^ ハンセン、ペル・ジュエル;ビョルンセン、ピーター・コーフォード。ハンセン、ベニ・ワインディング (1997)。 「動物プランクトンの捕食と成長: 体サイズ 2 ~ 2 μm の範囲内での拡大」。陸水学と海洋学。42 (4): 687–704。書誌コード:1997LimOc..42..687H。土井:10.4319/lo.1997.42.4.0687。多くの著者の調査結果を要約します。
- ^ abcdef クレイマー、ドナルド・L. (2001). 「採餌行動」.進化生態学. doi :10.1093/oso/9780195131543.003.0024. ISBN 978-0-19-513154-3。
- ^ ab グリフィス, デイヴィッド (1980年11月). 「採餌コストと相対的な獲物サイズ」.アメリカン・ナチュラリスト. 116 (5): 743– 752. Bibcode :1980ANat..116..743G. doi :10.1086/283666. JSTOR 2460632. S2CID 85094710.
- ^ Wetzel, Robert G.; Likens, Gene E. (2000). 「捕食者と被食者の相互作用」. Limnological Analyses . pp. 257– 262. doi :10.1007/978-1-4757-3250-4_17. ISBN 978-1-4419-3186-3。
- ^ abcd Pianka, Eric R. (2011).進化生態学(第7版(電子書籍)). Eric R. Pianka. pp. 78– 83.
- ^ マッカーサー、ロバート・H. (1984). 「消費者選択の経済学」.地理生態学:種の分布パターン. プリンストン大学出版局. pp. 59– 76. ISBN 978-0-691-02382-3。
- ^ abcd Bell 2012、4~5ページ
- ^ イーストマン、ルーカス・B.; ティール、マーティン (2015). 「甲殻類捕食者と腐肉食動物の採餌行動」. ティール、マーティン; ワトリング、レス (編).ライフスタイルと摂食生物学. オックスフォード大学出版局. pp. 535– 556. ISBN 978-0-19-979706-6。
- ^ Perry, Gad (1999年1月). 「探索モードの進化:生態学的観点と系統学的観点」. The American Naturalist . 153 (1): 98– 109. Bibcode :1999ANat..153...98P. doi :10.1086/303145. PMID 29578765. S2CID 4334462.
- ^ ベル 2012、69–188ページ
- ^ Gremillet, D.; Wilson, RP; Wanless, S.; Chater, T. (2000). 「マユグロアホウドリ、国際漁業、そしてパタゴニア棚」.海洋生態学進歩シリーズ. 195 : 69– 280. Bibcode :2000MEPS..195..269G. doi : 10.3354/meps195269 .
- ^ Charnov, Eric L. (1976). 「最適採餌、限界値定理」.理論個体群生物学. 9 (2): 129– 136. Bibcode :1976TPBio...9..129C. doi :10.1016/0040-5809(76)90040-x. PMID 1273796.
- ^ レイノルズ、アンディ(2015年9月)「最適採餌の束縛からレヴィウォーク研究を解放する」Physics of Life Reviews . 14 : 59–83 . Bibcode :2015PhLRv..14...59R. doi :10.1016/j.plrev.2015.03.002. PMID 25835600.
- ^ブキャナン、マーク(2008年6月5日)「生態学的モデリング:動物 の本質を映し出す数学的鏡」Nature 453 (7196): 714– 716. doi : 10.1038/453714a . PMID 18528368.
- ^ ウィリアムズ, アマンダ C.; フラックスマン, サミュエル M. (2012). 「捕食者は獲物の資源の質を評価できるか?」動物行動学83 ( 4): 883– 890. doi :10.1016/j.anbehav.2012.01.008. S2CID 53172079.
- ^ Scharf, Inon; Nulman, Einat; Ovadia, Ofer; Bouskila, Amos (2006年9月). 「異なる条件下における2つの競合する採餌モードの効率評価」. The American Naturalist . 168 (3): 350– 357. Bibcode :2006ANat..168..350S. doi :10.1086/506921. PMID 16947110. S2CID 13809116.
- ^ abcdefghijk Moore, Talia Y.; Biewener, Andrew A. (2015). 「逃げるか、出し抜くか:より広範な生態学的・進化学的文脈における生体力学的研究のモデルシステムとしての捕食者と被食者の相互作用」統合比較生物学55 ( 6): 1188–97 . doi : 10.1093/icb/icv074 . PMID 26117833.
- ^ ab deVries, MS; Murphy, EAK; Patek SN (2012). 「待ち伏せ型捕食者の攻撃メカニズム:槍で突くシャコ」. Journal of Experimental Biology . 215 (Pt 24): 4374– 4384. Bibcode :2012JExpB.215.4374D. doi : 10.1242/jeb.075317 . PMID 23175528.
- ^ 「クーガー」. Hinterland Who's Who .カナダ野生生物局およびカナダ野生生物連盟. 2007年5月18日時点のオリジナルよりアーカイブ。2007年5月22日閲覧。
- ^ 「カワカマス科(カワカマス類)」(PDF)インディアナ州魚類野生生物局. 2018年9月3日閲覧。
- ^ ブレイ、ダイアン. 「Eastern Frogfish, Batrachomoeus dubius」.オーストラリアの魚類. 2014年9月14日時点のオリジナルよりアーカイブ。 2014年9月14日閲覧。
- ^ “Trapdoor spiders”. BBC. 2010年12月3日時点のオリジナルよりアーカイブ。 2014年12月12日閲覧。
- ^ 「トラップドアスパイダー」アリゾナ・ソノラ砂漠博物館2014年. 2014年12月12日閲覧。
- ^ Gazda, SK; Connor, RC; Edgar, RK; Cox, F. (2005). 「フロリダ州シーダーキー沖における集団狩猟を行うバンドウイルカ(Tursiops truncatus)の役割分担と分業」Proceedings of the Royal Society . 272 (1559): 135– 140. doi :10.1098/rspb.2004.2937. PMC 1634948. PMID 15695203 .
- ^ Tyus, Harold M. (2011). 魚類の生態と保全. CRC Press. p. 233. ISBN 978-1-4398-9759-1。
- ^ Combes, SA; Salcedo, MK; Pandit, MM; Iwasaki, JM (2013). 「異なる種類の獲物を追うトンボの捕獲成功率と捕獲効率」.統合比較生物学. 53 (5): 787– 798. doi : 10.1093/icb/ict072 . PMID 23784698.
- ^ Hubel, Tatjana Y.; Myatt, Julia P.; Jordan, Neil R.; Dewhirst, Oliver P.; McNutt, J. Weldon; Wilson, Alan M. (2016年3月29日). 「アフリカの野生の犬における狩猟のエネルギーコストとリターン」. Nature Communications . 7 11034. doi :10.1038/ncomms11034. PMC 4820543. PMID 27023457. 移動
狩猟戦略は、一時的な加速、パワー、スピードを重視する極端な戦略から、獲物を疲労させて捕獲を容易にする粘り強さと持久力重視の極端な戦略まで多岐にわたる。犬と人間は、移動狩猟において、スピードや機動性よりも持久力に頼って勝利を収めると考えられている。
- ^ Goldbogen, JA; Calambokidis, J.; Shadwick, RE; Oleson, EM; McDonald, MA; Hildebrand, JA (2006). 「ナガスクジラにおける採餌潜水と突進摂食の運動学」. Journal of Experimental Biology . 209 (7): 1231– 1244. Bibcode :2006JExpB.209.1231G. doi : 10.1242/jeb.02135 . PMID 16547295. S2CID 17923052.
- ^ Sanders, Jon G.; Beichman, Annabel C.; Roman, Joe; Scott, Jarrod J.; Emerson, David; McCarthy, James J.; Girguis, Peter R. (2015). 「ヒゲクジラは肉食動物と草食動物の両方に類似した独自の腸内微生物叢を持つ」Nature Communications . 6 8285. Bibcode :2015NatCo...6.8285S. doi :10.1038/ncomms9285. PMC 4595633. PMID 26393325 .
- ^ Forbes, L. Scott (1989). 「獲物の防御と捕食者のハンドリング行動:危険な獲物仮説」. Oikos . 55 (2): 155– 158. Bibcode :1989Oikos..55..155F. doi :10.2307/3565418. JSTOR 3565418.
- ^ ab Lang, Stephen DJ; Farine, Damien R. (2017). 「社会的捕食戦略を研究するための多次元的枠組み」. Nature Ecology & Evolution . 1 (9): 1230– 1239. Bibcode :2017NatEE...1.1230L. doi :10.1038/s41559-017-0245-0. PMID 29046557.
- ^ MacNulty, Daniel R.; Tallian, Aimee; Stahler, Daniel R.; Smith, Douglas W. (2014年11月12日). Sueur, Cédric (編). 「オオカミによるバイソン狩りの成功における集団規模の影響」. PLOS ONE . 9 (11) e112884. Bibcode :2014PLoSO...9k2884M. doi : 10.1371/journal.pone.0112884 . PMC 4229308 . PMID 25389760.
- ^ Power, R. John; Shem Compion, RX (2009年4月). 「ボツワナ、チョベ国立公園サブティにおけるライオンによるゾウの捕食」.アフリカ動物学. 44 (1): 36– 44. doi :10.3377/004.044.0104.
- ^ ボーシャン 2012、7~12ページ
- ^ ドーソン, ジェームズ・W. (1988).アリゾナ州におけるハリスホークの協力的繁殖システム(修士論文). hdl :10150/276864.[ページが必要]
- ^ Vail, Alexander L.; Manica, Andrea; Bshary, Redouan (2013年4月23日). 「魚類の協働狩猟における指示的ジェスチャー」. Nature Communications . 4 (1): 1765. Bibcode :2013NatCo...4.1765V. doi : 10.1038/ncomms2781 . PMID 23612306.
- ^ Yong, Ed (2013年4月24日). 「ハタはジェスチャーを使ってウツボを狩猟チームに勧誘する」ナショナルジオグラフィック. 2018年9月17日時点のオリジナルよりアーカイブ。 2018年9月17日閲覧。
- ^ トフト、クラウス(プロデューサー)(2007年)「Killers in Eden」(DVDドキュメンタリー)オーストラリア放送協会。2009年8月12日時点のオリジナルよりアーカイブ。ISBN R-105732-9.
- ^ ab Bryce, Caleb M.; Wilmers, Christopher C.; Williams, Terrie M. (2017). 「大型捕食者と被食者のエネルギーと回避行動:ピューマ vs. 猟犬」. PeerJ . 5 e3701. doi : 10.7717/peerj.3701 . PMC 5563439. PMID 28828280 .
- ^ マリヤ・マジャー、クリスティーナ・ホルム、ヤエル・ルビン、トリネ・ビルデ (2018). 「社会性クモにおける協調的採餌は食物ニッチを拡大するが、資源をめぐる集団内競争を相殺しない」. Scientific Reports . 8 (1): 11828. Bibcode :2018NatSR...811828M. doi :10.1038/s41598-018-30199-x. PMC 6081395. PMID 30087391 .
- ^ “待ち伏せ捕食者”.シブリー自然センター. 2021年8月2日時点のオリジナルよりアーカイブ。 2018年9月17日閲覧。
- ^ Elbroch, L. Mark; Quigley, Howard (2016年7月10日). 「孤独性肉食動物における社会的相互作用」Current Zoology . 63 (4): 357– 362. doi :10.1093/cz/zow080. PMC 5804185 . PMID 29491995.
- ^ クエンクア・ダグラス(2017年10月11日)「孤独なピューマが昼食をとるマウンテンライオンに」ニューヨーク・タイムズ。 2018年9月17日閲覧。
- ^ フローレス、ダン(2016年)『コヨーテ・アメリカ:自然と超自然の歴史』ベーシックブックス、ISBN 978-0-465-05299-8。
- ^ Stow, Adam; Nyqvist, Marina J.; Gozlan, Rodolphe E.; Cucherousset, Julien; Britton, J. Robert (2012). 「孤独性捕食者における行動症候群は体の大きさと成長率に依存しない」. PLOS ONE . 7 (2) e31619. Bibcode :2012PLoSO...731619N. doi : 10.1371/journal.pone.0031619 . PMC 3282768. PMID 22363687 .
- ^ 「クモはどのように狩りをするのか?」アメリカ自然史博物館。2014年8月25日。 2018年9月5日閲覧。
- ^ ウェセロー, ロナルド・M.; ヘア, J. ダニエル (2009). 「捕食/捕食性昆虫」.昆虫百科事典(第2版). pp. 837– 839. doi :10.1016/B978-0-12-374144-8.00219-8. ISBN 978-0-12-374144-8。
- ^ Bar-Yam. 「捕食者と被食者の関係」. ニューイングランド複雑系研究所. 2018年9月7日閲覧。
- ^ abc 「Predator & Prey: Adaptations」(PDF) . ロイヤルサスカチュワン博物館. 2012年. オリジナル(PDF)から2018年4月3日時点のアーカイブ。 2018年4月19日閲覧。
- ^ Vermeij, Geerat J. (1993). 『進化とエスカレーション:生命の生態史』プリンストン大学出版局pp. 11 and passim. ISBN 978-0-691-00080-0。
- ^ Getz, WM (2011). 「バイオマス変換ウェブは消費者資源モデリングへの統一的なアプローチを提供する」. Ecology Letters . 14 (2): 113–24 . Bibcode :2011EcolL..14..113G. doi :10.1111/j.1461-0248.2010.01566.x. PMC 3032891. PMID 21199247 .
- ^ シドロヴィッチ、ヴァディム (2011). 脊椎動物捕食者被食者コミュニティの分析:ベラルーシの遷移的混合林地帯におけるヨーロッパ森林地帯の研究. Tesey. p. 426. ISBN 978-985-463-456-2。
- ^ アンジェリチ、フランチェスコ・M. (2015). 問題のある野生生物:学際的アプローチ. シュプリンガー. p. 160. ISBN 978-3-319-22246-2。
- ^ Hayward, MW; Henschel, P.; O'Brien, J.; Hofmeyr, M.; Balme, G.; Kerley, GIH (2006). 「ヒョウ(Panthera pardus)の獲物の好み」. Journal of Zoology . 270 (2): 298– 313. doi :10.1111/j.1469-7998.2006.00139.x.
- ^ Pulliam, H. Ronald (1974). 「最適食餌理論について」. The American Naturalist . 108 (959): 59– 74. Bibcode :1974ANat..108...59P. doi :10.1086/282885. S2CID 8420787.
- ^ Sih, Andrew; Christensen, Bent (2001). 「最適食事理論:いつ効果があり、いつ、なぜ失敗するのか?」.動物行動学. 61 (2): 379– 390. doi :10.1006/anbe.2000.1592. S2CID 44045919.
- ^ Sprules, W. Gary (1972). 「サイズ選択的捕食と食物競争が高高度動物プランクトン群集に及ぼす影響」.生態学. 53 (3): 375– 386. Bibcode :1972Ecol...53..375S. doi :10.2307/1934223. JSTOR 1934223.
- ^ Owen-Smith, Norman; Mills, MGL (2008). 「アフリカ大型哺乳類食物網における捕食者と被食者の大きさの関係」. Journal of Animal Ecology . 77 (1): 173– 183. Bibcode :2008JAnEc..77..173O. doi : 10.1111/j.1365-2656.2007.01314.x . hdl : 2263/9023 . PMID 18177336.
- ^ コット 1940、12~13ページ
- ^ Lloyd JE (1965). 「ホタルにおける攻撃的な擬態:ホタルのファム・ファタレス」. Science . 149 (3684): 653– 654. Bibcode :1965Sci...149..653L. doi :10.1126/science.149.3684.653. PMID 17747574. S2CID 39386614.
- ^ フォーブス、ピーター(2009年)『まばゆいばかりと欺瞞:模倣とカモフラージュ』イェール大学出版局、134頁。ISBN 978-0-300-17896-8。
- ^ Bester, Cathleen (2025年5月2日). 「Striated frogfish」.フロリダ博物館. フロリダ大学. 2025年7月12日閲覧。
- ^ Ruppert, Edward E.; Fox, Richard S.; Barnes, Robert D. (2004). 『無脊椎動物学』(第7版). Cengage Learning. pp. 153– 154. ISBN 978-81-315-0104-7。
- ^ Cetaruk, Edward W. (2005). 「ガラガラヘビとその他のガラガラヘビ科」. Brent, Jeffrey (編).集中治療毒性学:重篤中毒患者の診断と管理. Elsevier Health Sciences. p. 1075. ISBN 978-0-8151-4387-1。
- ^ バルセロウ、ドナルド・G. (2008). 『天然物質の医学毒性学:食品、菌類、薬草、植物、有毒動物』ワイリー社、1028ページ、ISBN 978-0-470-33557-4。
- ^ Li, Min; Fry, BG; Kini, R. Manjunatha (2005). 「卵のみの食事:マーブルウミヘビ(Aipysurus eydouxii)の毒素プロファイル変化と生態への影響」. Journal of Molecular Evolution . 60 (1): 81– 89. Bibcode :2005JMolE..60...81L. doi :10.1007/s00239-004-0138-0. PMID 15696370. S2CID 17572816.
- ^ Castello, ME; A. Rodriguez-Cattaneo; PA Aguilera; L. Iribarne; AC Pereira; AA Caputi (2009). 「弱電気魚 Gymnotus coropinae (Hoedeman) における波形生成:電気器官と電気器官放電」. Journal of Experimental Biology . 212 (9): 1351– 1364. Bibcode :2009JExpB.212.1351C. doi : 10.1242/jeb.022566 . PMID 19376956.
- ^ Feulner, PG; M. Plath; J. Engelmann; F. Kirschbaum; R. Tiedemann (2009). 「電気的な愛:電気魚は種特異的な放電を用いて交尾相手を認識する」. Biology Letters . 5 (2): 225– 228. doi :10.1098/rsbl.2008.0566. PMC 2665802. PMID 19033131 .
- ^ Catania, Kenneth C. (2015). 「電気ウナギは高電圧を用いて高速移動する獲物を追跡する」Nature Communications . 6 (1) 8638. Bibcode :2015NatCo...6.8638C. doi :10.1038/ncomms9638. PMC 4667699. PMID 26485580 .
- ^ Kramer, Bernd (1996).魚類における電気受容とコミュニケーション. 第42巻. レーゲンスブルク大学. doi :10.5283/epub.2108. ISBN 978-3-437-25038-5。
- ^ カラソフ, ウィリアム H.; ダイアモンド, ジャレッド M. (1988). 「消化における生理学と生態学の相互作用」.バイオサイエンス. 38 (9): 602– 611. doi :10.2307/1310825. JSTOR 1310825.
- ^ ラクストン、シェラット&スピード 2004、pp. vii–xii
- ^ カロ 2005, 67–114ページ
- ^ Merilaita, Sami; Scott-Samuel, Nicholas E.; Cuthill, Innes C. (2017年5月22日). 「カモフラージュの仕組み」. Philosophical Transactions of the Royal Society B: Biological Sciences . 372 (1724). doi :10.1098/rstb.2016.0341. PMC 5444062. PMID 28533458 .
- ^ カロ 2005、13~15ページ
- ^ Bergstrom, CT; Lachmann, M. (2001). 「警戒音は捕食者警戒のコストの高いシグナル:警戒心の強いバブラーのゲーム」.動物行動学. 61 (3): 535– 543. doi :10.1006/anbe.2000.1636. S2CID 2295026.
- ^ Getty, T. (2002). 「識別力のあるチメドリと最適な餌を求めるタカが出会う」.動物行動学. 63 (2): 397– 402. doi :10.1006/anbe.2001.1890. S2CID 53164940.
- ^ コット 1940、241–307ページ
- ^ Bowers, MD; Brown, Irene L.; Wheye, Darryl (1985). 「蝶の個体群における選択因子としての鳥類捕食」. Evolution . 39 (1): 93– 103. doi : 10.1111/j.1558-5646.1985.tb04082.x . PMID 28563638. S2CID 12031679.
- ^ Berenbaum, MR (1995年1月3日). 「防御の化学:理論と実践」.米国科学アカデミー紀要. 92 (1): 2– 8. Bibcode :1995PNAS...92....2B. doi : 10.1073/pnas.92.1.2 . PMC 42807. PMID 7816816 .
- ^ ラクストン、シェラット&スピード 2004年、70~81ページ
- ^ カロ 2005, 663–684ページ
- ^ ボーシャン 2012、83~88ページ
- ^ クラウス, イェンス; ラクストン, グレアム・D. (2002年10月10日). 『集団生活』 オックスフォード大学出版局. pp. 13– 15. ISBN 978-0-19-850818-2。
- ^ ラクストン、シェラット&スピード 2004年、54~55ページ
- ^ Dominey, Wallace J. (1983). 「コロニー性営巣魚類、特にブルーギル(Lepomis macrochirus)におけるモビング」Copeia . 1983 (4): 1086– 1088. doi :10.2307/1445113. JSTOR 1445113.
- ^ Brodie, Edmund D. (2009年11月3日). 「毒素と毒液」. Current Biology . 19 (20): R931 – R935 . Bibcode :2009CBio...19.R931B. doi : 10.1016/j.cub.2009.08.011 . PMID 19889364. S2CID 9744565.
- ^ コット 1940、368–389ページ
- ^ Merilaita, Sami; Vallin, Adrian; Kodandaramaiah, Ullasa; et al. (2011年7月26日). 「クジャクチョウにおける眼点の数と無知な捕食者に対する威嚇効果」.行動生態学. 22 (6): 1326– 1331. doi : 10.1093/beheco/arr135 .
- ^ Cumming, Jeffrey M.; Sinclair, Bradley J.; Triplehorn, Charles A.; Aldryhim, Yousif; Galante, Eduardo; Marcos-Garcia, Ma Angeles; Edmunds, Malcolm; Edmunds, Malcolm; Lounibos, L. Phillip; Frank, J. Howard; Showler, Allan T.; Yu, Simon J.; Capinera, John L.; Heppner, John B.; Philogène, Bernard JR; Lapointe, Stephen L.; Capinera, John L.; Capinera, John L.; Nayar, Jai K.; Goettel, Mark S.; Nation, James L.; Heppner, John B.; Negron, Jose F.; Heppner, John B.; Kondratieff, Boris C.; Schöning, Caspar; Stewart, Kenneth W.; Aldryhim, Yousif;ヘップナー, ジョン・B.; ハンゲイ, ジョージ (2008). 「デイマティック行動」.昆虫学百科事典. pp. 1173– 1174. doi :10.1007/978-1-4020-6359-6_863. ISBN 978-1-4020-6242-1。
- ^ Caro 2005、v–xi、4–5 ページ
- ^ カロ 2005, 413–414ページ
- ^ ジェイコブス&バスティアン 2017、4ページ
- ^ バルボサ、ペドロ、カステリャーノス、イグナシオ (2005).捕食者と被食者の相互作用の生態学. オックスフォード大学出版局. p. 78. ISBN 978-0-19-987454-5。
- ^ Janis, CM; Wilhelm, PB (1993). 「第三紀に哺乳類の追跡捕食者は存在したか? オオカミの化身とのダンス」. Journal of Mammalian Evolution . 1 (2): 103– 125. doi :10.1007/bf01041590. S2CID 22739360.
- ^ abc リチャード・ドーキンス、JR・クレブス(1979). 「種間および種内における軍備競争」Proceedings of the Royal Society B: Biological Sciences . 205 (1161): 489– 511. Bibcode :1979RSPSB.205..489D. doi :10.1098/rspb.1979.0081. PMID 42057. S2CID 9695900.
- ^ ab Abrams, Peter A. (1986年11月). 「捕食者の被捕食者への適応反応と被捕食者の捕食者への適応反応:軍拡競争アナロジーの失敗」. Evolution . 40 (6): 1229–1247 . doi : 10.1111/j.1558-5646.1986.tb05747.x . PMID 28563514. S2CID 27317468.
- ^ abcde Brodie, Edmund D. (1999年7月). 「捕食者と被食者の軍拡競争」. BioScience . 49 (7): 557– 568. doi : 10.2307/1313476 . JSTOR 1313476.
- ^ Vermeij, GJ (1994年11月). 「種間の進化的相互作用:選択、エスカレーション、そして共進化」. Annual Review of Ecology and Systematics . 25 (1): 219– 236. Bibcode :1994AnRES..25..219V. doi :10.1146/annurev.es.25.110194.001251.
- ^ ジェイコブス&バスティアン 2017、8ページ
- ^ ジェイコブス&バスティアン 2017、107ページ
- ^ Sheriff, Michael J.; Peacor, Scott D.; Hawlena, Dror; Thaker, Maria; Gaillard, Jean-Michel (2020). 「非消費的捕食者による被食者個体群サイズへの影響:証拠の不足」. Journal of Animal Ecology . 89 (6): 1302– 1316. Bibcode :2020JAnEc..89.1302S. doi : 10.1111/1365-2656.13213 . PMID 32215909.
- ^ Preisser, Evan L.; Bolnick, Daniel I.; Benard, Michael F. (2005). 「死ぬほど怖い? 捕食者と被食者の相互作用における威嚇と消費の影響」.生態学. 86 (2): 501– 509. Bibcode :2005Ecol...86..501P. doi :10.1890/04-0719. ISSN 0012-9658.
- ^ ab Peckarsky, Barbara L.; Abrams, Peter A.; Bolnick, Daniel I.; Dill, Lawrence M.; Grabowski, Jonathan H.; Luttbeg, Barney; Orrock, John L.; Peacor, Scott D.; Preisser, Evan L.; Schmitz, Oswald J.; Trussell, Geoffrey C. (2008年9月). 「古典を再考する:教科書的な捕食者と被食者の相互作用における非消費的影響の考察」. Ecology . 89 (9): 2416– 2425. Bibcode :2008Ecol...89.2416P. doi :10.1890/07-1131.1. PMID 18831163.
- ^ リンデマン、レイモンド・L. (1942). 「生態学の栄養動態的側面」.生態学. 23 (4): 399– 417. Bibcode :1942Ecol...23..399L. doi :10.2307/1930126. JSTOR 1930126.
- ^ Ordiz, Andrés; Bischof, Richard; Swenson, Jon E. (2013). 「大型肉食動物の保護は進むが、頂点捕食者は失われるのか?」. Biological Conservation . 168 : 128–133 . Bibcode :2013BCons.168..128O. doi : 10.1016/j.biocon.2013.09.024 . hdl : 11250/2492589 .
- ^ Pimm, SL; Lawton, JH (1978). 「複数の栄養段階における摂食について」. Nature . 275 (5680): 542– 544. Bibcode :1978Natur.275..542P. doi :10.1038/275542a0. S2CID 4161183.
- ^ Fedriani, Jose M.; Fuller, Todd K.; Sauvajot, Raymond M.; York, Eric C. (2000年10月). 「3種の同所性肉食動物における競争とギルド内捕食」. Oecologia . 125 (2): 258– 270. Bibcode :2000Oecol.125..258F. doi :10.1007/s004420000448. hdl : 10261/54628 . PMID: 24595837. S2CID : 24289407.
- ^ 「生態系におけるエネルギー伝達」ナショナルジオグラフィック2023年2月18日2023年2月18日閲覧。
- ^ Bond, WJ (2012). 「11. キーストーン種」. Schulze, Ernst-Detlef; Mooney, Harold A. (編). 『生物多様性と生態系機能』 . Springer. p. 237. ISBN 978-3-642-58001-7。
- ^ Botkin, D.; Keller, E. (2003).環境科学:生きている惑星としての地球. John Wiley & Sons. p. 2. ISBN 978-0-471-38914-9。
- ^ ab ウィリアム・J.リップル、ロバート・L.ベシュタ (2004).「オオカミと恐怖の生態学:捕食は生態系の構造を危険にさらすか?」バイオサイエンス. 54 (8): 755. doi : 10.1641/0006-3568(2004)054[0755:WATEOF]2.0.CO;2 .
- ^ ニール、ディック (2004).集団生物学入門. ケンブリッジ大学出版局. pp. 68– 69. ISBN 978-0-521-53223-5。
- ^ ネルソン, エリック H.; マシューズ, クリストファー E.; ローゼンハイム, ジェイ A. (2004年7月). 「捕食者は被食者の行動変化を誘導することで被食者の個体数増加を抑制する」(PDF) .生態学. 85 (7): 1853– 1858. Bibcode :2004Ecol...85.1853N. doi :10.1890/03-3109. JSTOR 3450359.
- ^ Krebs, Charles J.; Boonstra, Rudy; Boutin, Stan; Sinclair, ARE (2001). 「シラカワウサギの10年周期を駆動するものは何か?」BioScience . 51 (1): 25. doi : 10.1641/0006-3568(2001)051[0025:WDTYCO]2.0.CO;2 . hdl : 1807/359 .
- ^ クレブス、チャーリー、マイヤーズ、ジュディ(2014年7月12日)「スノーシューウサギの10年周期 ― 警告の物語」エコロジカル・ランツ。ブリティッシュコロンビア大学。 2018年10月2日閲覧。
- ^ 「捕食者とその獲物」BBC Bitesize . BBC . 2012年5月2日時点のオリジナルよりアーカイブ。 2015年10月7日閲覧。
- ^ Goel, Narendra S.; Maitra, SC; Montroll, EW (1971). 『相互作用する集団のボルテラモデルとその他の非線形モデルについて』Academic Press. ISBN 978-0-12-287450-5。
- ^ ab レビン, サイモン・A.; カーペンター, スティーブン・R.; ゴッドフレー, H. チャールズ・J.; キンジグ, アン・P.; ロロー, ミシェル; ロソス, ジョナサン・B.; ウォーカー, ブライアン; ウィルコーブ, デイビッド・S. (2009). 『プリンストン生態学ガイド』 プリンストン大学出版局. pp. 204–209. ISBN 978-1-4008-3302-3。
- ^ ウィリアム・W・マードック、シェリル・J・ブリッグス、ロジャー・M・ニスベット(2013年)『消費者資源ダイナミクス』プリンストン大学出版局、39頁。ISBN 978-1-4008-4725-9。
- ^ Nowak, Martin; May, Robert M. (2000).ウイルスダイナミクス:免疫学とウイルス学の数学的原理. オックスフォード大学出版局. p. 8. ISBN 978-0-19-158851-8。
- ^ Genovart, M.; Negre, N.; Tavecchia, G.; Bistuer, A.; Parpal, L.; Oro, D. (2010). 「若者、弱者、病人:捕食による自然淘汰の証拠」. PLOS ONE . 5 (3) e9774. Bibcode :2010PLoSO...5.9774G. doi : 10.1371/journal.pone.0009774 . PMC 2841644. PMID 20333305 .
- ^ ロックウッド 2009、281ページ
- ^ ロックウッド 2009、246ページ
- ^ ロックウッド 2009、271–272ページ
- ^ ロックウッド 2009、272~273ページ
- ^ ab Cushing, JM (2005年3月30日). 「書評:集団生物学における数学」アメリカ数学会報. 42 (4): 501– 506. doi : 10.1090/S0273-0979-05-01055-4 .
- ^ Thieme, Horst R. (2003). 人口生物学における数学. プリンストン大学出版局. ISBN 978-0-691-09291-1。
- ^ Kozlov, Vladimir; Vakulenko, Sergey (2013年7月3日). 「ロトカ・ヴォルテラ系におけるカオスについて:解析的アプローチ」.非線形性. 26 (8): 2299– 2314. Bibcode :2013Nonli..26.2299K. doi :10.1088/0951-7715/26/8/2299. S2CID 121559550.
- ^ Sih, Andrew (1987). 「被食者避難所と捕食者-被食者間の安定性」.理論個体群生物学. 31 (1): 1– 12. Bibcode :1987TPBio..31....1S. doi :10.1016/0040-5809(87)90019-0.
- ^ McNair, James N. (1986). 「避難所が捕食者-被食者相互作用に及ぼす影響:再考」.理論個体群生物学. 29 (1): 38– 63. Bibcode :1986TPBio..29...38M. doi :10.1016/0040-5809(86)90004-3. PMID 3961711.
- ^ ベリーマン、アラン・A.; ホーキンス、ブラッドフォード・A.; ホーキンス、ブラッドフォード・A. (2006). 「生態学と進化における統合概念としての避難所」オイコス. 115 (1): 192– 196.書誌コード:2006Oikos.115..192B. doi :10.1111/j.0030-1299.2006.15188.x.
- ^ クレスマン、ロス;ガライ、ヨーゼフ(2009)「捕食者と被食者の避難システム:生態系における進化的安定性」理論個体群生物学76 ( 4): 248–57 . Bibcode :2009TPBio..76..248C. doi :10.1016/j.tpb.2009.08.005. PMID 19751753.
- ^ Abrams, PA (2000). 「捕食者と被食者の相互作用の進化:理論と証拠」. Annual Review of Ecology and Systematics . 31 (1): 79– 105. Bibcode :2000AnRES..31...79A. doi :10.1146/annurev.ecolsys.31.1.79.
- ^ ab グリマルディ, デイヴィッド;エンゲル, マイケル・S. (2005).昆虫の進化. ケンブリッジ大学出版局. pp. 155–160. ISBN 978-0-521-82149-0。
- ^ Grant, SWF; Knoll, AH; Germs, GJB (1991). 「ナミビア、原生代後期ナマ層群における石灰化メタファーテスの可能性:起源、続成作用、そしてその意義」. Journal of Paleontology . 65 (1): 1– 18. Bibcode :1991JPal...65....1G. doi :10.1017/S002233600002014X. JSTOR 1305691. PMID 11538648. S2CID 26792772.
- ^ Awramik, SM (1971年11月19日). 「先カンブリア時代の柱状ストロマトライトの多様性:後生動物の出現を反映して」. Science . 174 (4011): 825– 827. Bibcode :1971Sci...174..825A. doi :10.1126/science.174.4011.825. PMID 17759393. S2CID 2302113.
- ^ スタンリー、スティーブン・M. (2008). 「海底における捕食が競争に打ち勝つ」.古生物学. 34 (1): 1– 21. Bibcode :2008Pbio...34....1S. doi :10.1666/07026.1. S2CID 83713101.
- ^ ロロン, コレンティン C.; レインバード, ロバート H.; ターナー, エリザベス C.; ワイルダー・グリーンマン, J.; ジャヴォー, エマニュエル J. (2018). 「真核生物の大進化における選択的捕食の影響:カナダ北極圏の証拠」.生命科学の新興トピックス. 2 (2): 247– 255. doi :10.1042/ETLS20170153. PMID 32412621. S2CID 92505644.
- ^ ab Dunn, FS; Kenchington, CG; Parry, LA; Clark, JW; Kendall, RS; Wilby, PR (2022年7月25日). 「英国チャーンウッドの森のエディアカラ紀から発見されたクラウングループの刺胞動物」. Nature Ecology & Evolution . 6 (8): 1095– 1104. Bibcode :2022NatEE...6.1095D. doi :10.1038/s41559-022-01807-x. PMC 9349040. PMID 35879540 .
- ^ ケリー、パトリシア(2003年)『化石記録における捕食者と被食者の相互作用』シュプリンガー、pp. 113– 139, 141–176および一部抜粋。ISBN 978-1-4615-0161-9. OCLC 840283264。
- ^ Daley, Allison C. (2013). 「アノマロカリス科」. Current Biology . 23 (19): R860 – R861 . Bibcode :2013CBio...23.R860D. doi : 10.1016/j.cub.2013.07.008 . PMID 24112975.
- ^ Anderson, PSL; Westneat, M. (2009). 「ダンクルオステウス・テレリ(節足動物、板皮類)の摂食運動の生体力学的モデル」.古生物学. 35 (2): 251– 269. Bibcode :2009Pbio...35..251A. doi :10.1666/08011.1. S2CID 86203770.
- ^ Carr, Robert K. (2010). 「ダンクルオステウス・テレリ(板皮類:節足動物)の古生態学」. Kirtlandia . 57 : 36–45 .
- ^ サンプソン, スコット・D.; ローウェン, マーク・A. (2005年6月27日). 「ユタ州上部白亜紀(マーストリヒチアン)ノースホーン層産のティラノサウルス・レックス:生物地理学的および古生態学的示唆」. Journal of Vertebrate Paleontology . 25 (2): 469– 472. doi :10.1671/0272-4634(2005)025[0469:TRFTUC]2.0.CO;2. S2CID 131583311.
- ^ Darimont, CT; Fox, CH; Bryan, HM; Reimchen, TE (2015年8月20日). 「ヒト捕食者の独特な生態」. Science . 349 (6250): 858– 860. Bibcode :2015Sci...349..858D. doi :10.1126/science.aac4249. PMID 26293961. S2CID 4985359.
- ^ ガブリエル、オットー、フォン・ブラント、アンドレス (2005).世界の魚類捕獲法. ブラックウェル. ISBN 978-0-85238-280-6。
- ^ グリフィン、エマ(2008年)『ブラッド・スポーツ:1066年以降のイギリスにおける狩猟』イェール大学出版局、ISBN 978-0-300-14545-8。
- ^ キング、リチャード・J.(2013年10月1日)『悪魔のウミウ:自然史』ニューハンプシャー大学出版局、9ページ。ISBN 978-1-61168-225-0。
- ^ グレイシャー、フィリップ(1998年)『鷹狩りとホーキング』バッツフォード、ISBN 978-0-7134-8407-6。
- ^ Aegerter, James; Fouracre, David; Smith, Graham C. (2017). Olsson, I Anna S. (編). 「英国におけるペットの猫と犬の個体群構造と密度の初推定」. PLOS ONE . 12 (4) e0174709. Bibcode :2017PLoSO..1274709A. doi : 10.1371/journal.pone.0174709 . PMC 5389805. PMID 28403172 .
- ^ 米国動物愛護協会「米国のペット所有統計」2012年4月27日閲覧。
- ^ リーベンバーグ、ルイス (2008). 「持続的狩猟と人類進化の関連性」.人類進化ジャーナル. 55 (6): 1156– 1159. Bibcode :2008JHumE..55.1156L. doi :10.1016/j.jhevol.2008.07.004. PMID 18760825.
- ^ 「Food For Thought」(PDF) .哺乳類の生活.英国放送協会. 2002年10月31日.
- ^ フリント、メアリー・ルイーズ; ドライシュタット、スティーブ・H. (1998). 『天敵ハンドブック:生物学的害虫防除の図解ガイド』 カリフォルニア大学出版局. ISBN 978-0-520-21801-7。[ページが必要]
- ^ ジョンストン、キース・M. (2013). 『SF映画:批評的入門』バーグ出版社98頁. ISBN 978-0-85785-056-0。
- ^ ニュービー、リチャード (2018年5月13日). 「『プレデター』はついに価値のある続編を手に入れるのか?」ハリウッド・レポーター. 2018年9月7日閲覧。
- ^ シャッツ、トーマス. 「ニューハリウッド」.映画ブロックバスターズ. p. 25.ジュリアン・ストリンガー著(2003年)『映画大ヒット作』ラウトレッジ社、 15~ 44頁 。ISBN 978-0-415-25608-7。
- ^ ピーター・デイヴィソン(2002年12月1日)「テッド・ヒューズ著『捕食者と獲物|選集 1957-1994』」ニューヨーク・タイムズ。 2018年10月5日閲覧。
ヒューズの初期の作品には、驚くほど多くの詩が収められていた。…魚や鳥、野獣や森の獣、捕食者と獲物の力強い化身。学生時代のヒューズは文学ではなく人類学を専攻し、詩を書き始める前に瞑想によってトランス状態のような前意識状態に入ることを選んだ。彼の詩は、初期にせよ後期にせよ、生き物との関係性に深く入り込み、動物の意識に近いところまで入り込んでいる。例えば、
「思考の狐」、
「エスターの雄猫」、
「パイク」など。
- ^ グールド、スティーブン・ジェイ(1995). 「歯と爪の100周年記念」.干し草の山の中の恐竜. ハーモニー・ブックス. pp. 63– 75. ISBN 978-0-517-70393-9。
- ^ abcdef Wallner, Astrid (2005年7月18日). 「神話における捕食者の役割」. WaldWissen森林管理情報. 2018年10月5日時点のオリジナルよりアーカイブ。 2018年10月5日閲覧。Wallner, A. (1998) Die Bedeutung der Raubtiere in der Mythologie: Ergebnisse einer Literaturstudieより翻訳。 – インフ.bl. Forsch.bereiches Landsch.ökol。 39:4-5。
- ^ ケラート, スティーブン・R.; ブラック, マシュー; ラッシュ, コリーン・リード; バス, アリスター・J. (1996). 「北米における人類文化と大型肉食動物の保全」.保全生物学. 10 (4): 977– 990. Bibcode :1996ConBi..10..977K. doi :10.1046/j.1523-1739.1996.10040977.x.
出典
- ボーシャン、ガイ(2012)『社会的捕食:集団生活が捕食者と被捕食者にもたらす利益』エルゼビアISBN 978-0-12-407654-9。
- Bell, WJ (2012).探索行動:資源発見の行動生態学. Springer Netherlands. ISBN 978-94-011-3098-1。
- カロ、ティム(2005年)『鳥類と哺乳類における捕食者防御』シカゴ大学出版局、ISBN 978-0-226-09436-6。
- コット、ヒュー・B. (1940).動物の適応的色彩. メシューエン. OCLC 974070031.
- ジェイコブス、デイビッド・スティーブ、バスティアン、アンナ(2017)『捕食者と被食者の相互作用:コウモリと被食者の共進化』シュプリンガー、ISBN 978-3-319-32492-0。
- ロックウッド、ラリー・L. (2009).個体群生態学入門. ジョン・ワイリー・アンド・サンズ. p. 281. ISBN 978-1-4443-0910-2。
- ラクストン, グレアム・D. ;シェラット, トム・N. ; スピード, マイケル・P. (2004). 『攻撃の回避:隠蔽、警告信号、擬態の進化生態学』 オックスフォード大学出版局. ISBN 978-0-19-852859-3。
{kind=link}
{kind=link}