トウモロコシ


| トウモロコシ | |
|---|---|
 | |
| 雄花と雌花を含む | |
| 科学的分類 | |
| 王国: | 植物界 |
| クレード: | 維管束植物 |
| クレード: | 被子植物 |
| クレード: | 単子葉植物 |
| クレード: | ツユクサ類 |
| 注文: | ポアレス |
| 家族: | イネ科 |
| 亜科: | パニコイデ科 |
| 属: | トウモロコシ |
| 種: | Z. mays |
| 二名法名 | |
| トウモロコシ | |

トウモロコシ(/ m eɪ z / ; Zea mays)は、北米英語ではcornとしても知られ、穀粒を生産する背の高い頑丈なイネ科植物です。この植物の葉のついた茎から、花粉を作る雄花序または房と、穂と呼ばれる雌花序が生じます。穂からは穀粒または種子と呼ばれる穀物が収穫されます。現代の商業用品種では、穂は通常黄色か白色ですが、他の品種には様々な色のものがあります。トウモロコシは、約 9,000 年前にメキシコ南部の先住民によって野生のテオシントから栽培化されました。ネイティブアメリカンは、スリーシスターズ複合栽培で豆やカボチャと一緒にトウモロコシを栽培しました。
トウモロコシの繁殖は人力に依存しています。コロンブス交換以来、トウモロコシは世界の多くの地域で主食となり、その総生産量は小麦や米を上回っています。トウモロコシの多くは、穀物(飼料)として、あるいは植物体全体(草姿)として家畜の飼料として利用されています。植物体は、飼料として梱包したり、より食味の良いサイレージに加工したりできます。スイートコーンと呼ばれる糖分の多い品種は食用として栽培され、畑で栽培されるトウモロコシ品種は家畜の飼料、コーンミール(マサ)、コーンスターチ、コーンシロップ、圧搾してコーン油、バーボンウイスキーなどのアルコール飲料、エタノールやその他のバイオ燃料などの化学原料として使用されます。
トウモロコシは世界中で栽培されており、毎年他のどの穀物よりも多くの重量が生産されています。2020年の世界生産量は11億トンでした。トウモロコシは多くの害虫や病気に悩まされており、中でもヨーロッパアワノメイガとコーンルートワームという2大害虫は、それぞれ米国で年間10億ドルの損失をもたらしています。現代の植物育種により、生産量と栄養価、干ばつ耐性、害虫や病気への耐性などの品質が大幅に向上しました。現在、多くのトウモロコシが遺伝子組み換えされています。
トウモロコシは、メキシコのトルティーヤやタマーレ、イタリアのポレンタ、アメリカのホミニーグリッツなど、様々な料理に使われています。トウモロコシのタンパク質には必須アミノ酸が不足しており、含まれるナイアシンはアルカリ処理によって遊離することでのみ利用可能になります。コロンブス以前のメソアメリカでは、トウモロコシはトウモロコシの神として神格化され、彫刻に描かれていました。
説明

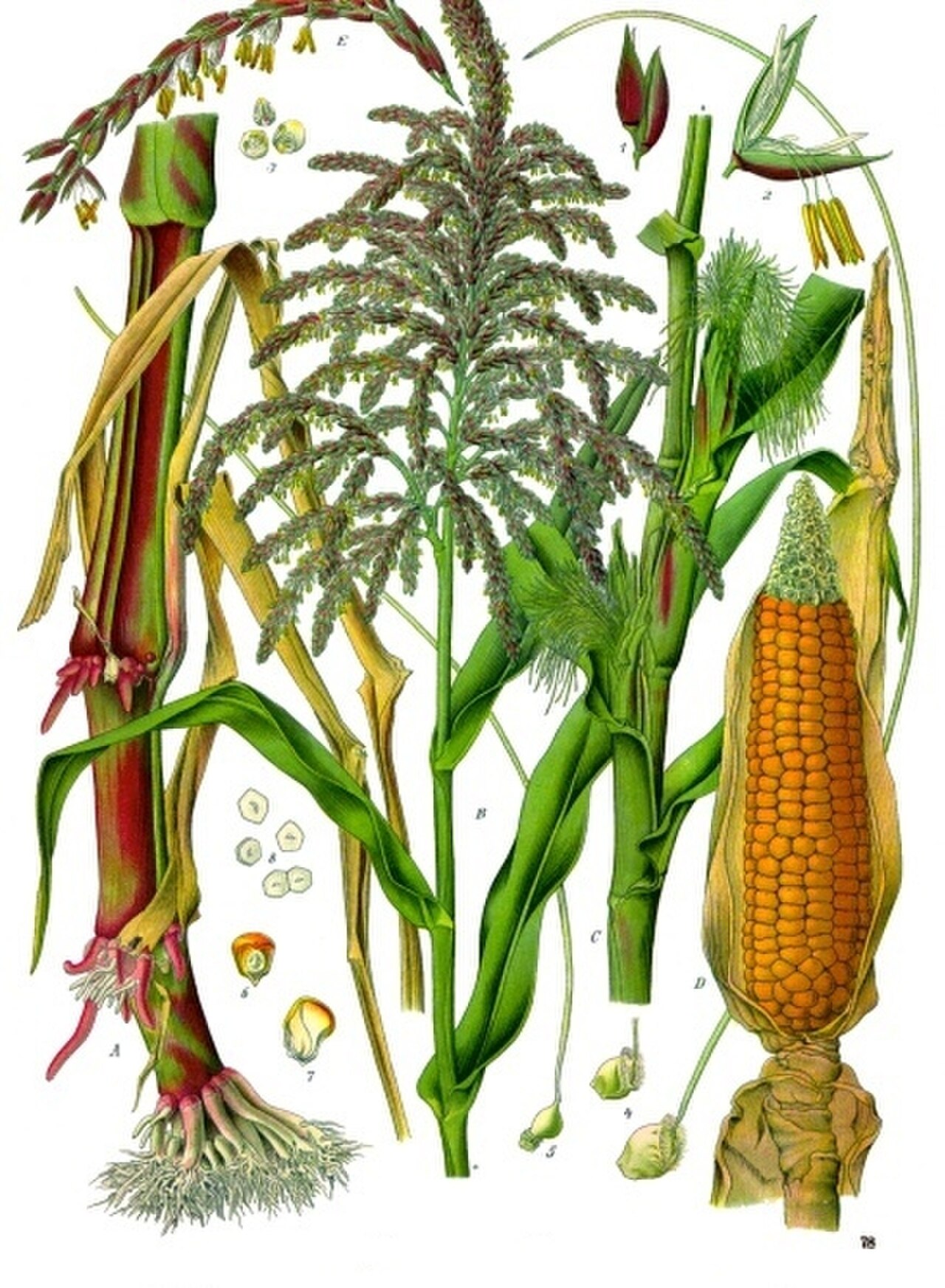
トウモロコシは、茎が 1 本しかない背の高い一年草で、高さは 1.2 〜 4 メートル (4 〜 13 フィート) です。[ 2 ]細長い葉は、節または節から、茎の反対側で交互に生じます。[ 2 ]トウモロコシは雌雄同株で、同じ植物に雄花と雌花が別々に咲きます。[ 2 ]茎の上部には雄花の花序である雄穂があります。雄蕊から花粉が放出され、風によって散布されます。[ 2 ]雄穂から茎を少し下ったところにある雌花の花序は、最初は絹糸、つまり柔らかい管状の毛の束として見られ、雌花ごとに心皮ごとに 1 本あり、これが種子 (種子と呼ばれることが多い。植物学的には、すべてのイネ科植物と同様に果実であり、種皮と融合して穎果を形成します)に成長して受粉します[ 3 ]。[ 2 ]雌花序全体が穂またはトウモロコシの芯に成長し、複数の葉の層または殻に包まれます。[ 2 ]穂葉は、成長中の穂に最も密接に関連する葉です。この葉とその上位の葉は、穀粒を構成する炭水化物(デンプン)の4分の3以上を供給します。 [ 4 ]
現代の品種では、穀粒は通常黄色または白色ですが、オレンジ色、赤色、茶色、青色、紫色、または黒色の品種もあります。穀粒は穂軸の周りに8列から32列に並んでおり、大きな穂軸には最大1200個の穀粒が付くこともあります。[ 5 ]黄色のトウモロコシはカロテノイドに由来し、赤色のトウモロコシはアントシアニンとフロバフェンに由来します。オレンジ色と緑色の品種には、これらの色素が組み合わさっている場合があります。[ 6 ]
トウモロコシは短日性光周性を有し、開花には一定時間以上の夜が必要である。さらに、開花には10℃(50℉)以上の暖かい日が十分必要である。開花の制御は遺伝的に決定されており、その生理学的メカニズムにはフィトクロム系が関与している。熱帯品種は、高緯度地域で栽培すると問題が生じる可能性がある。日照時間が長くなると、冬が来る前に種子を結実させる代わりに、植物が背丈以上に成長してしまう可能性があるからである。一方、急速に背丈が伸びることは、バイオ燃料の生産に都合が良い可能性がある。[ 2 ]
未熟なトウモロコシの芽には、強力な抗生物質物質である2,4-ジヒドロキシ-7-メトキシ-1,4-ベンゾキサジン-3-オン(DIMBOA)が蓄積され、広範囲の害虫に対するある程度の保護を提供します。[ 7 ]トウモロコシは根が浅いため、干ばつの影響を受けやすく、栄養分の乏しい土壌に耐えられず、強風によって根こそぎにされやすいです。[ 8 ]
花粉はアレルゲンですが、そのほとんどは穂から数メートル以内に落ち、リスクは主に農業従事者に限定されます。[ 9 ]
- たくさんの小さな雄花が集まって房を形成します。
- 若い絹糸を持つ雌花序
- 茎、穂、絹糸
- 完全に成長したトウモロコシ
- 茎についた成熟したトウモロコシの穂




.JPG/1280px-Klip_kukuruza_uzgojen_u_Me%C4%91imurju_(Croatia).JPG)
- 雄花
- 成熟したシルク


遺伝学

トウモロコシは20本の染色体を持つ二倍体です。ゲノム内の対立遺伝子変異の83%は、主にトウモロコシ属の種が交雑しやすい性質に由来する、テオシント系の祖先に由来しています。[ 10 ]バーバラ・マクリントックはトウモロコシを用いて「ジャンピング遺伝子」のトランスポゾン理論を検証し、1983年のノーベル生理学・医学賞を受賞しました。[ 11 ]トウモロコシは遺伝学と発生生物学において重要なモデル生物であり続けています。[ 12 ] MADSボックスモチーフはトウモロコシの花の発達に関与しています。[ 13 ]
トウモロコシ遺伝学・ゲノミクスデータベースは、トウモロコシ研究を支援するため、米国農務省(USDA)から資金提供を受けている。[ 14 ]国際トウモロコシ・小麦改良センターは、害虫抵抗性が検査されカタログ化されたトウモロコシ系統の大規模なコレクションを維持している。[ 15 ] 2005年に、米国国立科学財団、USDA、およびエネルギー省は、トウモロコシゲノムの配列を決定するコンソーシアムを結成した。結果として得られたDNA配列データは、ゲノム配列データの公共リポジトリであるGenBankに直ちに預けられた。 [ 16 ]トウモロコシゲノムの配列決定は2008年に完了した。[ 17 ] 2009年に、コンソーシアムは配列決定の取り組みの結果を発表した。[ 18 ]ゲノムの85%はトランスポゾンで構成され、32,540個の遺伝子が含まれている。その多くはトウモロコシのDNA内の転移因子のグループであるヘリトロンによって複製され、再編成されている。[ 19 ]
分類学
外部系統発生
トウモロコシ属(Zea)はソルガムと比較的近縁で、共に旧世界イネ科植物のPACMAD系統に属しています。一方、イネやコムギとははるかに遠縁で、これらはイネ科植物のもう一つの主要なグループであるBOP系統に属しています。また、ガマグラス( Tripsacum )とも近縁です。[ 20 ]
| (イネ科の一部 ) | |
トウモロコシとテオシント

トウモロコシは、その野生近縁種である4種のテオシントの栽培化変種である。[ 21 ]テオシント起源説は、1931年にロシアの植物学者ニコライ・イワノビッチ・ヴァヴィロフによって、 1932年にアメリカのノーベル賞受賞者ジョージ・ビードルによって提唱された。 [ 22 ]:10 2つの植物は外観が異なり、トウモロコシは1本の長い茎と複数の葉を持つのに対し、テオシントは短く茂った植物である。この2つの違いは、grassy tillers-1 ( gt1 , A0A317YEZ1 )とteosinte branched-1 ( tb1 , Q93WI2 )と呼ばれるわずか2つの遺伝子の違いによって大きく左右される。[ 21 ] 1930年代後半、ポール・マンゲルスドルフは、栽培トウモロコシは未知の野生トウモロコシと近縁属のトリプサカム属の種との交雑の結果であると示唆したが、これは現代の遺伝子検査によって反証されている。[ 22 ]
2004年、ジョン・ドーブリーは、メキシコ南西部の高地バルサス川流域原産のバルサス・テオシンテ(Zea mays subsp. parviglumis)を、現代のトウモロコシに遺伝的に最も類似した作物の野生近縁種であると特定した。 [ 23 ] [ 24 ]短いバルサス川流域の中央部は、初期の栽培化が行われた可能性が高い場所である。ゲレロ州イグアラ近郊の洞窟にある8,700年前の堆積層からは、トウモロコシの残留物が付いた石臼が発見されている。[ 25 ]ドーブリーとその同僚は2002年に、トウモロコシは約9,000年前に一度だけ栽培化され、その後アメリカ大陸全体に広がったことを示した。[ 26 ]
タバスコ州サンアンドレス島からカリブ海沿岸で、 7,300年前のトウモロコシの花粉が発見されました。 [ 25 ] 7,000年前、メキシコ南部、中央アメリカ、南アメリカ北部では原始的なトウモロコシが栽培されていました。オアハカ渓谷のギラ・ナキッツ洞窟で発見された初期のトウモロコシの穂の考古学的遺跡は、およそ6,250年前のものです。プエブラ州テワカン近郊の洞窟で発見された最古の穂は、5,450年前のものです。[ 27 ]
北へ広がる
約4,500年前、トウモロコシは北方へと広がり始めました。トウモロコシは、約4,100年前、ニューメキシコ州とアリゾナ州のいくつかの場所で初めて栽培されました。[ 27 ]西暦1千年紀には、トウモロコシの栽培は北部の地域でより広く普及しました。特に、北米東部でトウモロコシの大規模な農業と消費が始まったのは、西暦900年頃です。ネイティブアメリカンは、この新しい作物のために広大な森林と草原を切り開きました。[ 28 ] 500年から1,000年前、現在の米国南東部でトウモロコシ栽培が増加したのと時を同じくして、環境の変化に非常に敏感な淡水産二枚貝が減少しました。 [ 29 ]
名前
トウモロコシという名称は、タイノ語のmahisのスペイン語形maízに由来する。[ 30 ]スウェーデンの植物学者カール・リンネは、Zea maysの種小名として一般名 maize を使用した。[ 31 ] maizeという名称は、文脈や地理的な地域によって変わる複雑で多様な意味を持つcornとは異なり、この 1 つの穀物だけを指すため、正式用法、科学的用法、国際的な用法において一般名として好まれている。 [ 32 ]ほとんどの国では主にmaizeという用語を使用し、 cornという名称は主に米国と他の英語圏の国で使用されている。[ 33 ] [ 34 ]トウモロコシという用語を主に使用する国では、 cornという単語はあらゆる穀物作物を示す可能性があり、地理的には地元の主食によって異なる。[ 35 ]例えば、イングランドでは小麦、スコットランドやアイルランドではオート麦など。[ 32 ]トウモロコシをトウモロコシと呼ぶようになったのは、18世紀の北米で「インディアンコーン」を短縮したのが始まりである。 [ 36 ]
食の歴史家ベティ・ファッセルは、北米における「トウモロコシ」という言葉の歴史に関する記事の中で、 「トウモロコシという言葉を口にすることは、言語と歴史における悲劇的な茶番劇的な誤訳に陥ることだ」と述べている。[ 37 ]イギリスでの使用法と同様に、スペイン人もトウモロコシを穀物の総称である「パニーソ」と呼び、イタリア人も「ポレンタ」と呼んでいた。後にイギリス人はトウモロコシを「トルコ小麦」、「トルココーン」、「インディアンコーン」と呼ぶようになった。ファッセルは「彼らは場所ではなく状態、文明化された穀物ではなく未開の穀物を意味していた」と述べている。[ 37 ]
国際農業生物科学センターなどの国際団体は、トウモロコシを好ましい一般名とみなしている。[38] トウモロコシという言葉は、国連食糧農業機関[ 39 ] 、メキシコ国際トウモロコシ・小麦改良センター、インドトウモロコシ研究所[40]、オーストラリアトウモロコシ協会[ 41]、ナイジェリア国立トウモロコシ協会[42]、ガーナ国立トウモロコシ協会[ 43 ] 、南アフリカトウモロコシトラスト[ 44 ] 、ジンバブエ種子トウモロコシ協会[ 45 ]などで使用されている。
栽培
コロンブス以前の発展

トウモロコシの繁殖には人の手が必要である。自然繁殖する祖先種であるテオシントの穀粒は自然に穂軸から落ちるが、栽培化されたトウモロコシの穀粒は落ちない。[ 46 ]すべてのトウモロコシは約9,000年前のメキシコ南部における単一の栽培化から生まれた。現存する最古のトウモロコシの種類はメキシコ高地のものである。トウモロコシはこの地域から低地へ、そしてアメリカ大陸へ2つの主要な経路で広がった。[ 26 ]栽培化の中心はおそらくメキシコ中南部のバルサス川渓谷であった。 [ 47 ]トウモロコシは少なくとも8,000年前にはエクアドル高地に到達した。[ 48 ] 7,600年前までに中央アメリカ南部に到達し、7,000年から6,000年前の間にコロンビアのアンデス山脈の渓谷に到達した。[ 47 ]
最も初期のトウモロコシは、1株につき小さな穂が1本だけ生えていました。[ 5 ]オルメカ人とマヤ人はメソアメリカ全土で様々な品種のトウモロコシを栽培し、調理、粉砕、ニシュタマリゼーションによる加工を行っていました。[ 27 ] 3000年前までに、トウモロコシはオルメカ文化の中心となり、暦、言語、神話にも影響を与えました。[ 37 ]
チリ中南部のマプチェ族は、スペイン到来以前の時代にキヌアやジャガイモとともにトウモロコシを栽培していた。 [ 49 ]インカ帝国の拡大以前は、トウモロコシはチリからアンデス山脈を越えて運ばれ、アルゼンチンのラカル県メリンキナの南緯40度まで取引・輸送されていた。[ 50 ]
コロンブス交換

1492年にヨーロッパ人が到着すると、スペイン人入植者はトウモロコシを消費し、探検家や貿易商がそれをヨーロッパに持ち帰りました。スペイン人入植者はトウモロコシよりも小麦パンを好みました。当時のキリスト教信仰では、小麦だけが聖体変化を起こしてキリストの体へと変化できると考えられていたため、聖餐のパンにおいてトウモロコシ粉を小麦の代わりに使用することはできませんでした。[ 51 ]
トウモロコシは多様な気候条件で生育できることから、世界中に広まりました。コロンブスの航海のわずか数十年後にはスペインで栽培され、その後イタリア、西アフリカ、フィリピンなどにも広がりました。[ 51 ] [ 52 ] 17世紀には、南ヨーロッパの農民の一般的な食料となりました。18世紀には、南フランスとイタリアの農民の主食となり、特にイタリアではポレンタとして食べられました。 [ 53 ]
トウモロコシが西洋の農業システムに導入されたとき、その生産性の高さから歓迎された。しかし、トウモロコシが主食となった地域では、すぐに栄養失調が蔓延する問題が生じた。[ 54 ]アメリカ先住民は 少なくとも紀元前1200年から1500年ごろから、灰と石灰で作ったアルカリ水にトウモロコシを浸す方法を習得しており、ニシュタマリゼーションという工程を生み出した。彼らはトウモロコシの殻を取り除くためにこれを行ったが、偶然にもビタミンB群のナイアシンも同時に放出され、ナイアシンの欠乏がペラグラの原因となった。[ 55 ]アルカリ処理と食事の多様性が理解され、適用されるようになってから、ペラグラは先進国から姿を消した。リジンを多く含むトウモロコシの開発と、よりバランスの取れた食事の推進が、ペラグラの減少に貢献した。ペラグラは、食糧が乏しい地域や難民キャンプで、寄付されたトウモロコシで人々が生き延びているところでは、今もなお存在している。[ 56 ]
従来の育種
先史時代のトウモロコシの品種改良は、大きな穂をつける大型の植物を生み出した。近代の品種改良は、個人が自らの畑で生産性の高い品種を選び、その種子を他の農家に販売することから始まった。ジェームズ・L・リードは、最も初期かつ最も成功した品種改良者の一人であり、1860年代にリードのイエロー・デントを開発した。これらの初期の取り組みは、集団選択(一列の植物を一方の親の種子から育てる)と受粉後の植物の選択(雌親のみが判明する)に基づいていた。その後の品種改良の取り組みには、穂から列への選択(CG・ホプキンス、1896年頃)、選抜された近交系から作られた交配種(GH・シュル、1909年)、そして4つの近交系を使った非常に成功した二重交配種( DF・ジョーンズ、1918年頃、1922年)などがあった。大学が支援する品種改良プログラムは、近代の交配種の開発と導入において特に重要であった。[ 57 ]
1940年代以降、トウモロコシの最良の品種は、収量、栄養、干ばつ、害虫・病害耐性といった特定の特性が最適化された近親交配種から作られた第一世代のハイブリッド種でした。従来の交配と遺伝子組み換えの両方が、生産量の増加と耕作地、農薬、水、肥料の必要性の削減に成功しました。トウモロコシの収量ポテンシャルが過去数十年で増加したという仮説を裏付ける矛盾する証拠があります。これは、収量ポテンシャルの変化は、個々の植物あたりの収量ポテンシャルの増加ではなく、葉の角度、倒伏抵抗性、高密度植栽への耐性、病害・害虫耐性、その他の農業特性に関連していることを示唆しています。[ 58 ]
トウモロコシの特定の品種は、穂を多く生産するように品種改良されており、アジア料理で野菜として使われる「ベビーコーン」の原料となっている。[ 59 ] [ 60 ]ミニトウモロコシと呼ばれる開花の早い品種は、1年間で複数世代が得られるため、科学研究に役立てるために開発された。[ 61 ]オロトンと呼ばれる品種は、窒素固定微生物との共生関係を進化させ、植物の窒素の29%~82%を供給している。[ 62 ]国際トウモロコシ・小麦改良センター(CIMMYT)は、最適化された品種を提供するための従来の育種プログラムを運営している。このプログラムは1980年代に開始された。[ 63 ]ハイブリッド種子は、アフリカ向けの干ばつ耐性トウモロコシプロジェクトによってアフリカに配布されている。[ 64 ]
熱帯在来種は、病害抵抗性および草食動物抵抗性の両方において、重要でありながら十分に活用されていない抵抗性アレルの供給源であり続けている。このようなアレルは、生産性の高い品種に導入することができる。 [ 65 ]この目的のための希少アレルは、Dao氏とSood氏によって2014年に発見された。[ 65 ] 2018年、CIMMYTのZerka Rashid氏は、熱帯の干ばつ耐性形質のために開発された関連マッピングパネルを用いて、ソルガムべと病抵抗性をもたらす新たなゲノム領域を発見し、既知の差次的メチル化領域をさらに特徴づけた。[ 66 ]
遺伝子工学
遺伝子組み換えトウモロコシは、 2016年に商業的に栽培された26種類の遺伝子組み換え作物のうちの1つでした。 [ 67 ] [ 68 ]このうちの大部分はBtトウモロコシです。遺伝子組み換えトウモロコシは1997年以降、米国とカナダで栽培されており、[ 69 ] 2016年までに米国のトウモロコシ作物の92%が遺伝子組み換えでした。[ 67 ] 2011年時点で、除草剤耐性トウモロコシと害虫耐性トウモロコシの品種はそれぞれ20カ国以上で栽培されていました。[ 70 ]
2000年9月、動物の食用としてのみ承認されていたスターリンク遺伝子組み換えトウモロコシが含まれていたため、最大5000万ドル相当の食品がリコールされました。 [ 71 ]
成長中
トウモロコシは耐寒性が低いため、温帯地域では春に植えなければなりません。根系は一般的に浅いため、土壌水分に依存します。C4炭素固定を利用する植物であるトウモロコシは、アルファルファや大豆などのC3炭素固定を利用する植物よりも、はるかに水効率の高い作物です。トウモロコシは、花が受粉の準備を整える絹糸出芽期に干ばつの影響を受けやすいです。アメリカ合衆国では、伝統的にトウモロコシが「7月4日までに膝の高さまで成長」していれば豊作と予測されていましたが、現代のハイブリッド種は一般的にこの成長率を超えています。サイレージ用のトウモロコシは、植物が緑色で果実が未熟なうちに収穫されます。スイートコーンは、受粉後、デンプンが形成される前の「ミルクステージ」、つまり晩夏から初秋から中秋にかけて収穫されます。畑で栽培されるトウモロコシは、穀粒を完全に乾燥させるため、秋の終わり頃まで畑に放置されます。実際には、冬、あるいは早春まで収穫されないこともあります。十分な土壌水分の重要性は、アフリカの多くの地域で実証されています。そこでは、定期的な干ばつによってトウモロコシの不作とそれに伴う飢饉が頻繁に発生しています。トウモロコシは主に湿潤で暑い気候で栽培されますが、寒冷、暑熱、乾燥、湿潤といった条件でも生育できるため、非常に用途の広い作物です。[ 72 ]
トウモロコシは、アメリカ先住民によって、スリーシスターズと呼ばれる複合栽培システムで、小さな丘陵地帯に植えられました。[ 73 ]トウモロコシは豆類の生育を支え、豆類は、豆類やその他のマメ科植物の根に生息する窒素固定根粒菌から得られる窒素を供給し、カボチャは土壌を覆うことで雑草を防ぎ、蒸発を防ぐ地被植物となりました。[ 74 ]
- 播種から3週間後の苗
- 若い茎
- 成熟した植物は穂を出す
- 確立された広大な土地でのトウモロコシ種子作物




収穫
スイートコーンは、穀物用トウモロコシよりも早く収穫され、品種によって60日から100日で成熟します。乳熟期に収穫されるスイートコーンの収穫期間を延長するには、成熟の早い品種と遅い品種を選抜するか、2週間ごとに異なる場所に植え付けることで対応できます。[ 75 ] 穀物用トウモロコシは、収穫時期を迎えてから比較的長期間、場合によっては数ヶ月間も圃場で保存できます。乾燥した状態で保管すれば、殻付きのまま収穫・貯蔵できます。[ 76 ]
米国農務省によると、1855年から1894年までの40年間で、1ブッシェルのトウモロコシを生産するために必要な労働時間は4時間34分からわずか41分に短縮されました。[ 77 ] 1940年より前、北米のトウモロコシは大半が手作業で収穫されていました。これには多数の労働者とそれに伴う社交行事(蜂の殻むき)が含まれていました。1850年代以降、1列または2列の機械式ピッカー(穂を摘み取り、茎葉は残す)やトウモロコシ専用の刈り取り機であるコーンバインダーなど、工程を部分的に機械化する機械が登場しました。後者はショックを与えることができる束を作ります。手作業または機械式ピッカーによって穂全体が収穫されますが、穂から穀粒を取り除くにはトウモロコシ脱穀機の別の操作が必要です。トウモロコシの穂は、しばしばトウモロコシ倉庫に丸ごと保管され、一部の家畜の飼料として十分でした。今日では、穂が丸ごと入ったトウモロコシ倉庫やトウモロコシバインダーはあまり一般的ではありません。なぜなら、ほとんどの近代的な農場はコンバインで畑から穀物を収穫し、それを貯蔵庫に保管するからです。トウモロコシヘッド(リールの代わりにポイントとスナップロール付き)付きのコンバインは茎を切断せず、単に茎を引き下げます。茎は下方に進み、地面にくしゃくしゃに折りたたまれて山になり、通常は土壌の有機物になるために残されます。トウモロコシの穂は大きすぎてプレートのスロットの間を通れません。スナップロールが茎を引き離すため、機械に入るのは穂と殻だけです。コンバインは殻と芯を分離し、粒だけを残します。[ 78 ]
- アイオワ州でのトウモロコシの収穫
- フィンランドのトウモロコシの収穫
- ミャンマーのトウモロコシの手摘み
- マラウイでトウモロコシの殻を手で剥く



.jpg/1280px-Africa_Food_Security_11_(10665081134).jpg)
穀物貯蔵庫
乾燥は、穀物をマイコトキシンで汚染するカビ菌による被害を防ぐ、あるいは少なくとも軽減するために不可欠です。アスペルギルス属とフザリウム属は最も一般的なマイコトキシン源であり、農業において重要な役割を担っています。[ 69 ]収穫した穀物の水分含有量が高すぎる場合は、穀物乾燥機を用いて加熱空気を穀物に吹き込むことで水分含有量を減らします。この作業には、可燃性ガス(プロパンガスまたは天然ガス)と送風機の駆動電力という形で大量のエネルギーが必要になる場合があります。 [ 79 ]
生産
トウモロコシは世界中で広く栽培されており、毎年他のどの穀物よりも多くの重量が生産されています。[ 80 ] 2020年の世界の総生産量は11億6000万トンで、米国が全体の31.0%を占めてトップを占めています(表)。中国は世界総生産量の22.4%を占めています。[ 81 ]
| トウモロコシのトップ生産者 | |
|---|---|
| 2020年 | |
| 数値(百万トン) | |
1.アメリカ合衆国 | 360.3 ( 31%) |
2.中国 | 260.7 ( 22.43%) |
3.ブラジル | 104 ( 8.95%) |
4.アルゼンチン | 58.4 ( 5.02%) |
5.ウクライナ | 30.3 ( 2.61%) |
6.インド | 30.2 (2.6%) |
7.メキシコ | 27.4 ( 2.36%) |
8.インドネシア | 22.5 (1.94%) |
9.南アフリカ | 15.3 ( 1.32%) |
10.ロシア | 13.9 (1.2%) |
| 世界合計 | 1162.4 |
| 出典:FAOSTAT [ 82 ] [ 81 ] | |
 トウモロコシの生産量(2023年)
トウモロコシの生産量(2023年)- トウモロコシ(ピンクの帯)はサトウキビに次いで2番目に広く生産されている主要作物であり、穀物作物の中では最も広く生産されています。[ 83 ]
.svg)
![トウモロコシ(ピンクの帯)はサトウキビに次いで2番目に広く生産されている主要作物であり、穀物作物の中では最も広く生産されています。[83]](/wiki/File:World_production_of_primary_crops_by_main_commodities.svg)
害虫

無脊椎動物、雑草、病原体など多くの害虫がトウモロコシの成長と発育に影響を与える可能性があります。 [ 84 ] [ 85 ]
トウモロコシは、多数の真菌性、細菌性、ウイルス性植物病にかかりやすい。経済的に重要な病気には、葉の病気、トウモロコシ黒穂病、穂腐れ病、茎腐れ病などがある。[ 86 ]トウモロコシ北部葉枯れ病は、その分布域全体でトウモロコシに被害を与え、縞状葉枯れ病と紋枯れ病はアジアで問題となっている。[ 87 ] [ 88 ]トウモロコシの真菌性病気の中には、アフラトキシンなどの潜在的に危険なマイコトキシンを生成するものがある。[ 69 ]アメリカ合衆国では、主要な病気にはタール斑点病、条斑細菌病、灰色葉斑点病、トウモロコシ北部葉枯れ病、ゴス萎凋病などがあり、2022年にはタール斑点病が最も被害が大きく、1億1,680万ブッシェルの損失をもたらした。[ 89 ]
米国ではトウモロコシは、ヨーロッパアワノメイガ(ECB、Ostrinia nubilalis)とコーンルートワーム(Diabrotica spp) 、ウェスタンコーンルートワーム、ノーザンコーンルートワーム、サザンコーンルートワームの2大害虫によって、それぞれ年間10億ドル相当の損失を被っています。[ 90 ] [ 91 ] [ 92 ]もう1つの深刻な害虫は、ヨトウガ(Spodoptera frugiperda)です。[ 93 ] コクゾウムシ(Sitophilus zeamais)は、貯蔵穀物の深刻な害虫です。[ 94 ]ノーザンヨトウガ、オリエンタルヨトウガ、またはイネ耳切りイモムシ(Mythimna separata )は、アジアのトウモロコシの大きな害虫です。[ 95
線虫もトウモロコシの害虫です。すべてのトウモロコシには何らかの線虫が寄生している可能性があり、根におけるネグサレセンチュウの個体数は「膨大」になることがあります。特に水ストレスと雑草の防除が不十分な場合、植物への悪影響として、畑全体または部分的に発育不良が見られます。[ 96 ]
多くの植物、例えばイネ科の単子葉植物(イネ科)であるEchinochloa crus-galli(ヒエ科)や、アカザ科の双子葉植物(広葉草本)であるAmaranthus(アマランサス科)は、トウモロコシと競合し、作物の収量を減少させる可能性があります。防除には、機械による雑草除去、火炎除草、または除草剤の使用などが挙げられます。[ 97 ]
- ヨーロッパアワノメイガによるトウモロコシの芯の被害
- トウモロコシ畑のヨーロッパアワノメイガの幼虫

.JPG/1280px-Symptoms_corncobs_destruction_caused_by_Ostrinia_nubilalis_(cropped).JPG)
.jpg/1280px-European_Corn_Borer_(15350098570).jpg)
用途
料理
トウモロコシとコーンミール(乾燥させたトウモロコシの粉末)は、世界の多くの地域で主食となっています。 [ 5 ]トウモロコシは食品原料であるコーンスターチの製造に使用されます。[ 98 ]トウモロコシデンプンは加水分解され、酵素処理されて、甘味料である高果糖コーンシロップが製造されます。[ 99 ]トウモロコシは発酵・蒸留されてバーボンウイスキーが製造されます。[ 100 ] トウモロコシの胚芽からはコーン油が抽出されます。 [ 101 ]
先史時代、メソアメリカの女性はトウモロコシを挽いてコーンミールを作りました。陶器の容器が発明された後、オルメカ人はトウモロコシを豆と一緒に調理するようになり、主食の栄養価を高めました。トウモロコシには重要な栄養素であるナイアシンが天然に含まれていますが、ニシュタマル化のプロセスなしでは生体内に吸収されません。マヤ人はニシュタマル粉を使って粥やタマーレを作りました。[ 102 ]トウモロコシはメキシコ料理 の主食です。マサ(ニシュタマル)はトルティーヤ、アトーレ、その他多くの中米料理の主材料です。トウモロコシのトルティーヤ、タマーレ、アトーレ、そしてこれらをベースにした料理の主材料です。[ 103 ]トウモロコシに生える トウモロコシ黒穂病菌はウイトラコチェとして知られ、メキシコの珍味です。[ 104 ]
粗挽きのトウモロコシ粉は、多くの文化圏でとろみのある粥に加工されます。イタリアのポレンタ、ブラジルのアングー、ルーマニアのママリガ、アメリカのコーンミールマッシュ(アメリカ南部ではホミニーグリッツ)、南アフリカのミエリパップ、アフリカの他の地域ではサザ、ンシマ、ウガリなどと呼ばれる食品などです。16世紀にポルトガル人によってアフリカに導入されたトウモロコシは、アフリカで最も重要な主食作物となりました。[ 105 ]
スイートコーンは、糖分が多くデンプン質が少ない遺伝的品種で、未熟な状態で穂軸付きとうもろこしとして食べられます。[ 106 ]

.jpg/1280px-Summer_corn_(48286638996).jpg)


栄養価
| 100g(3.5オンス)あたりの栄養価 | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| エネルギー | 360 kJ (86 kcal) | ||||||||||||||||||||||||||||||||||||||||||||
18.7グラム | |||||||||||||||||||||||||||||||||||||||||||||
| スターチ | 5.7グラム | ||||||||||||||||||||||||||||||||||||||||||||
| 糖類 | 6.26グラム | ||||||||||||||||||||||||||||||||||||||||||||
| 食物繊維 | 2グラム | ||||||||||||||||||||||||||||||||||||||||||||
1.35グラム | |||||||||||||||||||||||||||||||||||||||||||||
3.27グラム | |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| その他の構成要素 | 量 | ||||||||||||||||||||||||||||||||||||||||||||
| 水 | 75.96グラム | ||||||||||||||||||||||||||||||||||||||||||||
USDA データベース エントリへのリンク中型サイズ (長さ 6-3/4 インチから 7-1/2 インチ) のトウモロコシの穂 1 本には、90 グラムの種子が入っています。 | |||||||||||||||||||||||||||||||||||||||||||||
| †成人に対する米国の推奨事項に基づいて推定された割合。 [ 107 ]ただし、カリウムについては米国アカデミーの専門家の推奨に基づいて推定されています。[ 108 ] | |||||||||||||||||||||||||||||||||||||||||||||
生の黄色い甘いトウモロコシの粒は、水分76%、炭水化物19% 、タンパク質3% 、脂肪1%で構成されています。100グラムのトウモロコシの粒は86カロリーで、ビタミンB群、チアミン、ナイアシン(遊離型の場合)、パントテン酸(B5) 、葉酸の優れた供給源です( 1日あたりの摂取量の10~19% )。[ 109 ]トウモロコシには必須アミノ酸のトリプトファンとリジンが最適量未満しか含まれていないため、タンパク質源としてはあまり適していません。[ 110 ]豆類やマメ科植物のタンパク質は、トウモロコシのタンパク質を補完します。[ 110 ]
動物飼料
トウモロコシは、動物飼料の主要な供給源である。穀物作物として、乾燥した穀粒は飼料として使用される。穀粒は、穂軸をつけたままトウモロコシ貯蔵庫に保管されることが多いが、殻をむいて穀物貯蔵庫に保管されることもある。穀粒を飼料に使用した場合、植物の残りの部分(茎葉)は、後で飼料、敷料、または土壌改良剤などとして使用できる。植物全体(穀粒と茎と葉)が飼料として使用される場合は、通常、細かく刻んでサイレージにする。これは、乾燥した形よりも反芻動物が消化しやすく、口当たりが良いためである。[ 111 ]伝統的に、トウモロコシは収穫後にショック(塊)に集められ、そこでさらに乾燥された。その後、家畜に与えるまで何ヶ月も貯蔵することができた。サイレージは、サイロまたはサイレージラッパーで製造することができる。熱帯地方では、トウモロコシは年間を通じて収穫され、動物の青刈飼料として与えられる。[ 112 ]俵に詰めたトウモロコシの茎は、家畜の飼料として干し草の代替品として利用されるほか、この目的のために栽培されたトウモロコシを直接放牧することもできます。 [ 113 ]
- トラックがトウモロコシの副産物でできた穀物飼料を飼い葉桶に撒いている間、牛たちは柵の横で待機している。
- 俵詰めされたトウモロコシの茎


化学薬品
トウモロコシのでんぷんは、プラスチック、布地、接着剤、その他多くの化学製品に加工することができます。[ 114 ]トウモロコシの湿式製粉工程で得られる水分を多く含む副産物であるコーンスティープリカーは、生化学産業や研究において、微生物を培養するための培地として使用されています。[ 115 ]
バイオ燃料
飼料用トウモロコシは暖房に利用されている。専用のトウモロコシストーブ(薪ストーブに似ている)は、飼料用トウモロコシか木質ペレットを使って熱を発生させる。トウモロコシの穂軸はバイオマス燃料源として利用できる。トウモロコシの粒を燃料として使う家庭用暖房炉には、粒を火に投入する大きなホッパーが付いている。[ 116 ]トウモロコシはエタノール燃料を生産するための原料として使われている。[ 117 ]食料価格は、バイオ燃料生産のためのトウモロコシの使用によって間接的に影響を受ける。つまり、そのような使用は需要を増加させ、したがってトウモロコシの価格も上昇させる。[ 118 ]オーストリア、ブルゲンラント州ストレムの先駆的なバイオマスガス化発電所が2005年に稼働を開始した。フィッシャー・トロプシュ法でバイオガスからディーゼルを生成することが可能である。[ 119 ]
- 2007年、ドイツ、ノイミュンスター近郊の農場に設置されたトウモロコシサイレージ消化槽。トウモロコシの実だけでなく、植物体全体を使用しています。緑色の防水シートで覆われた上部カバーは、消化槽内に貯蔵されたバイオガスによって支えられています。

人間の文化において
メソアメリカでは、トウモロコシは生命力の源とされ、トウモロコシの神(通常は女性)として神格化されている。[ 120 ]米国では、米国議会議事堂の柱頭にトウモロコシの穂が彫られている。[ 121 ]サウスダコタ州ミッチェルのコーンパレスでは、色とりどりのトウモロコシの穂軸と穂を使って、毎年リサイクルされる壁画のデザインが採用されている。[ 122 ]オハイオ州ダブリンのコンクリート製の「トウモロコシ畑」の彫刻には、草原に生えた何百ものトウモロコシの穂が描かれている。[ 123 ] 1993年以降に鋳造されたクロアチアの1リパ硬貨の裏面には、2つの熟した穂が付いたトウモロコシの茎が描かれている。 [ 124 ]
トウモロコシの粒は、臆病さを象徴することがある。トウモロコシは鶏の餌として使われるが、鶏は臆病者の象徴である。 1973年のチリクーデターの数ヶ月前、反アジェンデ派の抗議者たちは、アジェンデの退陣を求めて軍の兵舎にトウモロコシを投げつけた。[ 125 ] [ 126 ]
- メキシコ、カンペチェのマヤのトウモロコシの神の漆喰の頭部、西暦550~850年
- ジャイナ島の陶器で作られた、トウモロコシの穂から現れる若いマヤのトウモロコシ神の小像(西暦600~900年)
- トウモロコシを貯蔵するアステカ人、フィレンツェ写本、1540/1585年





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.JPG){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
.JPG){kind=link}
.jpg){kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
参照
- ブルーコーン – トウモロコシの品種
- デントコーン – トウモロコシの品種
- 雄花除去 – トウモロコシの雄花を取り除く
- フリントコーン – トウモロコシの品種
- グラスジェムコーン – トウモロコシの品種
- 収穫後の損失(穀物) - 穀物の損失が発生する仕組みと問題への対処方法
- 紫トウモロコシ – フリントトウモロコシの変種
- プッシュ・プル技術- トウモロコシとソルガムの害虫防除戦略
- ゼイン – トウモロコシに含まれるタンパク質
参考文献
- ^コントレラス、A.;ルイス・コラル、JA;メンジヴァル、J.アラゴン・クエバス、F.ゴンザレス・レデスマ、M.サンチェス、JJ (2019)。「ゼアメイズ」。IUCN 絶滅危惧種レッドリスト 2019: E.T77726273A77726310。土井: 10.2305/IUCN.UK.2019-2.RLTS.T77726273A77726310.en。
- ^ a b c d e f g Solaimalai, A.; Anantharaju, P.; Irulandi, S.; Theradimani, M. (2020年5月10日). 「6. 成長と発達の段階」 .トウモロコシ作物:改良、生産、保護、収穫後技術. CRC Press . ISBN 978-1-000-17695-7。
- ^ 「Caryopsis」 . Merriam-Webster . 2024年1月9日閲覧。
- ^ 「トウモロコシに殺菌剤を散布する前に:立ち止まって!見て!考えよう!」統合作物管理。アイオワ州立大学エクステンション。2021年7月24日閲覧。
- ^ a b cデイビッドソン、アラン (2014). 「トウモロコシ」.オックスフォード・コンパニオン・トゥ・フード(第3版).オックスフォード大学出版局. pp. 484– 486. ISBN 978-0-19-967733-7。
- ^ Chatham, Laura A.; Paulsmeyer, Michael; Juvik, John A. (2019). 「経済的な天然着色料の展望:トウモロコシからの洞察」.理論・応用遺伝学. 132 (11): 2927– 2946, 図1. doi : 10.1007/s00122-019-03414-0 . PMID 31451836. S2CID 201729476 .
- ^ Smith, C. Michael; Clement, Stephen L. (2012). 「植物の節足動物に対する抵抗性の分子基盤」Annual Review of Entomology . 57 (1): 309– 328. doi : 10.1146/annurev-ento-120710-100642 . PMID 21910639 .
- ^ 「トウモロコシの茎の倒伏」(PDF)モンサント・イマジン、2008年10月2日。2009年2月25日時点のオリジナル(PDF)からアーカイブ。 2009年2月23日閲覧。
- ^ Oldenburg, Marcus; Petersen, Arnd; Baur, Xaver (2011). 「トウモロコシ花粉は職業上曝露を受けた労働者にとって重要なアレルゲンである」 . Journal of Occupational Medicine and Toxicology . 6 (1): 32. doi : 10.1186 /1745-6673-6-32 . PMC 3269392. PMID 22165847 .
- ^ Wani, Shabir Hussain; Samantara, Kajal; Razzaq, Ali; Kakani, Grihalakshmi; Kumar, Pardeep (2022年6月). 「野生への回帰:高度な育種ツールを用いたトウモロコシ(Zea mays L.)の病害抵抗性の解明」. Molecular Biology Reports . 49 (6): 5787– 5803. doi : 10.1007/s11033-021-06815-x . PMID 35064401. S2CID 254834535 .
- ^ブラウン、デイビッド(2009年11月20日)「科学者たちはトウモロコシゲノムに大きな期待を寄せている」ワシントン・ポスト
- ^ストラブル、ジョシュ;スキャンロン、マイケル・J. (2009). 「トウモロコシ(Zea mays):植物生物学における基礎研究および応用研究のためのモデル生物」.コールド・スプリング・ハーバー・プロトコル. 2009 (10) pdb.emo132. doi : 10.1101/pdb.emo132 . ISSN 1940-3402 . PMID 20147033 .
- ^フリードマン、ウィリアム E.ムーア、リチャード C.プルガナン、マイケル D. (2004 年 10 月)。「植物発達の進化」。アメリカ植物学雑誌。91 (10)。ジョン・ワイリー&サンズ:1726–1741。ビブコード:2004AmJB...91.1726F。土井:10.3732/ajb.91.10.1726。PMID 21652320。アメリカ植物協会。
- ^ 「MaizeGDBへようこそ」MaizeGDB . 2024年1月11日閲覧。
- ^プラサンナ、ボッドゥパリ M.;ブルース、アナニ。ベイネ、ヨセフ。マクンビ、ダン。ゴウダ、マンジェ。アシム、ムハンマド。マルティネリ、サミュエル。責任者、グラハム P.パリミ、スリニバス(2022年11月)。「トウモロコシにおけるヨトウヨトウ管理における宿主植物の抵抗性:アフリカとアジアにおける関連性、現状および展望」。理論的および応用遺伝学。135 (11): 3897–3916。土井: 10.1007/s00122-022-04073-4。PMC 9729323。PMID 35320376。
- ^ “Welcome to MaizeSequence.org” . MaizeSequence.org. 2013年9月27日時点のオリジナルよりアーカイブ。 2024年3月12日閲覧。
- ^ 「研究者ら、主要作物トウモロコシのゲノムを解読」ロイター通信2008年2月26日2014年10月6日閲覧。
- ^ Schnable, PS; Ware, D.; Fulton, RS; Stein, JC; Wei, F.; et al. (2009). 「B73トウモロコシゲノム:複雑性、多様性、そしてダイナミクス」 . Science . 326 (5956): 1112– 1115. Bibcode : 2009Sci...326.1112S . doi : 10.1126/science.11 78534. PMID 19965430. S2CID 21433160 .
- ^ Feschotte, C.; Pritham, E. (2009). 「ヘリトロンの豊富な集合体がトウモロコシゲノムを形作る」 . Proceedings of the National Academy of Sciences . 106 (47): 19747– 19748. Bibcode : 2009PNAS..10619747F . doi : 10.1073/ pnas.0910273106 . PMC 2785235. PMID 19926864 .
- ^ Gaut, Brandon S.; Le Thierry d'Ennequin, Maud; Peek, Andrew S.; Sawkins, Mark C. (2000年6月20日). 「植物核ゲノムの進化モデルとしてのトウモロコシ」 . Proceedings of the National Academy of Sciences . 97 (13): 7008– 7015. Bibcode : 2000PNAS...97.7008G . doi : 10.1073/pnas.97.13.7008 . PMC 34377. PMID 10860964 .
- ^ a b Whipple, Clinton J.; Kebrom, Tesfamichael H.; Weber, Allison L.; Yang, Fang; Hall, Darren; et al. (2011年8月16日). 「grassy tillers1はトウモロコシの頂芽優勢を促進し、イネ科植物における日陰シグナルに反応する」 . Proceedings of the National Academy of Sciences . 108 (33): E506-12. doi : 10.1073/pnas.1102819108 . PMC 3158142. PMID 21808030 .
- ^ a bウィルクス、ギャリソン(2004年3月8日)。「第1章 1 トウモロコシ、奇妙で素晴らしい:しかし、明確な起源は分かっているのか?」スミス、C・ウェイン、ベトラン、ハビエル、ルンゲ、ECA(編)『トウモロコシ:起源、歴史、技術、生産』ジョン・ワイリー・アンド・サンズ、pp . 3-63。ISBN 978-0-471-41184-0。
- ^ Doebley, John F. (2004). 「トウモロコシの進化の遺伝学」(PDF) . Annual Review of Genetics . 38 : 37– 59. doi : 10.1146/annurev.genet.38.072902.092425 . PMID 15568971.オリジナル(PDF)から2010年6月12日時点のアーカイブ。 2010年5月30日閲覧。
- ^ Wu, Chi-Chih; Diggle, Pamela K.; Friedman, William E. (2011年9月). 「Balsas teosinte, Zea mays subsp. parviglumis (Poaceae)における雌配偶体の発育と重複受精」.有性植物生殖. 24 (3): 219– 229. doi : 10.1007/s00497-011-0164-1 . PMID 21380710. S2CID 8045294 .
- ^ a b Ranere, Anthony J.; Piperno, Dolores R.; Holst, Irene; Dickau, Ruth; Iriarte, José (2009). 「メキシコ、バルサス川中央渓谷における完新世初期におけるトウモロコシとカボチャの栽培化の文化的・年代的背景」 . Proceedings of the National Academy of Sciences . 106 (13): 5014– 5018. Bibcode : 2009PNAS..106.5014R . doi : 10.1073/pnas.0812590106 . PMC 2664064. PMID 19307573 . Ranere, Anthony J.; Piperno, Dolores R.; Holst, Irene; Dickau, Ruth; Iriarte, José (2009). 「メキシコ、バルサス川中央渓谷産、9千年紀初頭のトウモロコシのデンプン粒と植物珪酸体による証拠」 . Proceedings of the National Academy of Sciences . 106 (13): 5019– 5024. Bibcode : 2009PNAS..106.5019P . doi : 10.1073/ pnas.0812525106 . PMC 2664021. PMID 19307570 .
- ^ a b Matsuoka, Y.; Vigouroux, Y.; Goodman, MM; et al. (2002). 「多座マイクロサテライト遺伝子型解析によるトウモロコシの単一栽培化の証明」 . Proceedings of the National Academy of Sciences . 99 (9): 6080–4 . Bibcode : 2002PNAS...99.6080M . doi : 10.1073/pnas.052125199 . PMC 122905. PMID 11983901 .
- ^ a b cジョン・ロニー(2009年冬)「トウモロコシ農業の始まり」『アーキオロジー・サウスウェスト』23 (1):4.
- ^エマーソン, トーマス・E.; ヘッドマン, クリスティン・M.; サイモン, メアリー・L. (2005). 「限界的な園芸家かトウモロコシ農民か? ランフォード伝統のトウモロコシ消費に関する考古植物学的、古病理学的、同位体学的証拠」.ミッドコンチネンタル考古学ジャーナル. 30 (1): 67– 118. doi : 10.1179/mca.2005.003 . JSTOR 20708222. S2CID 129150225 .
- ^ピーコック, エヴァン; ハーグ, ウェンデル R.; ウォーレン, メルビン L. Jr. (2005). 「トウモロコシ農業の到来と一致する淡水ムール貝の先史時代の衰退」(PDF) .コンサベーション・バイオロジー. 19 (2): 547– 551. Bibcode : 2005ConBi..19..547P . doi : 10.1111/j.1523-1739.2005.00036.x . S2CID 3679709 .
- ^ 「maize, n. (and adj.)」オックスフォード英語辞典(オンライン版)。オックスフォード大学出版局。(サブスクリプションまたは参加機関のメンバーシップが必要です。)
- ^ラナム、ピーター;ペーニャ・ロサス、ファン・パブロ。ガルシア=カザル、マリア・ニエベス(2014年4月)。「世界のトウモロコシの生産、利用、消費」。ニューヨーク科学アカデミーの年報。1312 (1): 105–112。Bibcode : 2014NYASA1312..105R。土井:10.1111/nyas.12396。PMID 24650320。S2CID 4640742。
- ^ a bエンスミンガー、オードリー・H. (1994).食品と栄養百科事典、第2版. CRCプレス. p. 479. ISBN 978-0-8493-8980-1国際的には「maize」という語が好んで用いられます。
なぜなら、アメリカ合衆国でこの植物の名称として知られている「corn」という語は、多くの国で主要穀物と同義語となっているからです。例えば、イギリスでは「corn」は小麦を指し、スコットランドとアイルランドではオート麦を指します。
- ^マクレラン・プレイステッド、スーザン (2013). 「トウモロコシ」. アンドリュー・スミス編. 『オックスフォード・エンサイクロペディア・オブ・フード・アンド・ドリンク・イン・アメリカ(第2版)』. ニューヨーク:オックスフォード大学出版局. ISBN 978-0-19-973922-62023年2月15日閲覧。
他の多くの国で「トウモロコシ」と呼ばれているものに「コーン」という語を使うのは、アメリカ合衆国特有のものです。「小麦トウモロコシ」「大麦トウモロコシ」「ライ麦トウモロコシ」といった小粒の穀物に慣れていたヨーロッパ人は、アメリカ固有のトウモロコシを「インディアンコーン」と呼んでいました。この語は「コーン」に短縮され、アメリカ原産の植物を表す英語の単語となりました。
- ^エスピノーザ、マウリシオ (2015 年 4 月 1 日)。」「『トウモロコシはどれも同じ』、そしてアメリカの農作物の王様に関するその他の愚行」オハイオ州立大学:食品・農業・環境科学部。2020年12月3日時点のオリジナルよりアーカイブ。 2022年9月21日閲覧。
- ^ "corn, n.1" .オックスフォード英語辞典(オンライン版). オックスフォード大学出版局.(サブスクリプションまたは参加機関のメンバーシップが必要です。)
- ^メンケン, HL (1984). 『アメリカ言語:アメリカ合衆国における英語の発達に関する探究』(第4版). ニューヨーク:アルフレッド・A・クノップフ. p. 122. ISBN 0-394-40075-5正統派英語における「コーン」は、
人間の食用穀物、特に小麦を意味します(例えば、穀物法)。初期の入植者たちはこの用法に従い、スペイン人がインディアンに倣って「maiz(メイズ)」と呼んでいたものに「インディアンコーン」という名称を与えました。…しかし、この形容詞は徐々に使われなくなり、18世紀半ばには「maize(メイズ)」は単に「corn(コーン)」と呼ばれ、穀物全般は「breadstuff(パン類)」と呼ばれるようになりました。1774年、トーマス・ハッチンソンはジョージ3世に説教した際、「ライ麦とトウモロコシの混合物」という限定的な意味で「コーン」を使用しました。「どんなトウモロコシですか?」とジョージが尋ねました。ハッチンソンは「インディアンコーンです」と説明しました。「あるいは、文献でよく言われるように、maize(メイズ)」です。」
- ^ a b c Fussell, Betty (1999). 「トウモロコシをコーンに翻訳する:アメリカ在来穀物の変容」. Social Research . 66 (1): 41– 65. JSTOR 40971301. Gale A54668866 ProQuest 209670587. 「コーン」という言葉を口にすることは、言語と歴史における悲劇的かつ茶番的な誤訳に陥ることです。もしイギリス人がコロンブスに倣い、アラワク族が主食穀物と名付けたタイノ語「mahiz」を
音声化していたら、私たちは今日のような言語的窮地に陥ることはなかったでしょう。聖書のルツが「異国のトウモロコシの中で涙を流して立っていた」時、彼女は小麦畑に立っていたのだと、私は毎年誰かに説明しなければならないのです。しかし、コロンブスの航海日誌には「インディアンがmaizと呼んでいた穀物を…スペイン人はpanizoと呼んでいた」と記されており、スペイン人にとってもそれはほぼ同義語だった。スペイン語の「panizo」は、彼らが知っていた穀物――小麦、キビ、大麦、オート麦――の総称であり、イタリア語の「polenta」(ラテン語のpubに由来)も同様だった。英語の「corn」も同様で、「コンビーフ」のように塩の粒も含め、あらゆる種類の穀物を包括していた。
1536年、パリの植物学者を介したフランスの言語帝国主義によって「Turcicum frumentum」という用語が生まれ、イギリス人はこれをすぐに「トルコ小麦」「トルコトウモロコシ」「インディアントウモロコシ」と翻訳した。トルコやインディアンとは、場所ではなく状態、つまり文明化された穀物ではなく未開の穀物を指し、トルコ人もこれに同意し、「kukuruz」(野蛮なという意味)と呼んだ。
- ^ 「Zea mays (トウモロコシ)」 . CABIコンペンディウム. CABIコンペンディウム.国際農業・生物科学センター. 2019. doi : 10.1079/cabicompendium.57417 . 2022年9月16日閲覧。
- ^ 「トウモロコシ」国連食糧農業機関2022年9月16日閲覧。
- ^ 「概要 – ICAR-インドトウモロコシ研究所」 。 2022年10月5日時点のオリジナルよりアーカイブ。 2022年9月16日閲覧。
- ^ 「Maize Association - Maize Association Australia」 . 2022年9月16日閲覧。
- ^ 「ナイジェリアトウモロコシ協会、国の農業を支援したIITAを表彰」国際熱帯農業研究所。2022年9月16日閲覧。
- ^ 「SARD-SCトウモロコシ部門がガーナトウモロコシ協会の設立を支援」国際熱帯農業研究所(IATA)2016年3月18日。 2024年3月10日閲覧。
- ^デュ・プレシス、レオン. 「THE MAIZE TRUST:トウモロコシ産業の管理者」 . Grain SA . 2024年3月10日閲覧。
- ^ルシケ, ジョセフ; ドノヴァン, フィリップ A (1995). 「ジンバブエのトウモロコシ種子産業」.南部アフリカ開発. 12 (2): 189– 196. doi : 10.1080/03768359508439804 . ISSN 0376-835X .
- ^ Benz, BF (2001). 「オアハカ州ギラ・ナキッツ産テオシントの栽培化に関する考古学的証拠」 . Proceedings of the National Academy of Sciences . 98 (4): 2104– 2106. Bibcode : 2001PNAS...98.2104B . doi : 10.1073 / pnas.98.4.2104 . ISSN 0027-8424 . PMC 29389. PMID 11172083 .
- ^ a b Piperno, Dolores R. (2011年10月). 「新世界熱帯における植物栽培と家畜化の起源:パターン、プロセス、そして新たな展開」 . Current Anthropology . 52 (S4): S453– S470. doi : 10.1086/659998 . S2CID 83061925.トウモロコシの起源地とされるメキシコの中央バルサス川流域における最近の研究では
、8700年前のトウモロコシの珪酸体とデンプン粒の存在が確認されており、これはこの作物の記録上最も古い年代である(Piperno et al. 2009; Ranere et al. 2009)。膨大なデータ集積によると、トウモロコシは7600年前までに中央アメリカ南部に拡散し、7000年から6000年前の間にコロンビアのアンデス山脈間渓谷にまで移動していたことが示唆されています。コロンビアのカウカ渓谷には、初期のトウモロコシの存在を示す遺跡が数多く存在するため、トウモロコシが南アメリカに入ってからは、アンデス山脈間渓谷が主要な拡散経路であった可能性が高いと考えられます。
- ^パガン=ヒメネス、ハイメ R.;グアチャミン・テロ、アナ・M.ロメロ・バスティダス、マーサ・E。コンスタンティン・カストロ、アンジェロ・R. (2016 年 6 月)。 「9000年紀後半、エクアドル高原のクビラン地域でゼア・メイズ・L.が使用されていたことが古代のデンプンによって明らかになった」。第四次インターナショナル。404 : 137–155。Bibcode : 2016QuInt.404..137P。土井:10.1016/j.quaint.2015.08.025。
- ^ Dillehay, Tom D. ; Pino Quivira, Mario ; Bonzani, Renée ; Silva, Claudia ; Wallner, Johannes ; Le Quesne, Carlos (2007). 「チリ中南部における耕作湿地と新たな複雑性、そして気候変動の長距離影響」(PDF) . Antiquity . 81 (314): 949– 960. doi : 10.1017/s0003598x00096034 . S2CID 59480757 .
- ^ペレス、アルベルト E.;エラ、ジョージナ (2011)。「Identificación de maiz de vasijas recuperadas de la Patagonia noroccidental argentina」 [アルゼンチン、パタゴニア北西部の陶器の容器に残ったトウモロコシの識別]。マガッラニア(スペイン語)。39 (2): 309–316。土井: 10.4067/S0718-22442011000200022。hdl : 11336/42613。
- ^ a bアール、レベッカ(2012年)『征服者の身体:1492年から1700年にかけてのスペイン領アメリカにおける食糧、人種、植民地時代の経験』ケンブリッジ大学出版局、pp. 17, 144, 151。
- ^サラザール、AM;パスクアル、CB。モンタナ州カシリット;ペンテコステス、ケンタッキー州。デュマラグ、ペンシルバニア州;バージニア州ラディアジュニア;パリル、JF (2016)。「フィリピンの伝統的なトウモロコシ品種の育種の可能性」(PDF)。SABRAO 育種と遺伝学のジャーナル。48 (2): 154–161 .
{{cite journal}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ランガー、ウィリアム・L. (1975). 「アメリカの食料とヨーロッパの人口増加 1750–1850」.社会史ジャーナル. 8 (2): 51– 66. doi : 10.1353/jsh/8.2.51 . JSTOR 3786266 .
- ^ 「トウモロコシの起源:ペラグラの謎」。『食品を理解する』欧州食品情報評議会。2001年12月。2006年9月27日時点のオリジナルよりアーカイブ。 2006年9月14日閲覧。
- ^スタラー、ジョン、カラスコ、マイケル(2009年11月24日) 『プレ・コロンブス時代の食文化:古代メソアメリカにおける食、文化、市場への学際的アプローチ』シュプリンガー・サイエンス&ビジネス・メディア、317頁。ISBN 978-1-4419-0471-3。
- ^トンプソン, ジャニス・J.; マノーレ, メリンダ; ヴォーン, リンダ (2016年1月15日). 「エネルギー代謝に関与する栄養素」 . 『栄養学の科学』 .ピアソン・エデュケーション. pp. 292– 321. ISBN 978-0-13-429880-1。
- ^ Jugenheimer, Robert W. (1958). 「農業開発論文 #62」.ハイブリッドトウモロコシの育種と種子生産. ローマ:国連食糧農業機関.
- ^ Duvick, DN; Cassman, KG (2009). 「緑の革命後、米国中北部における温帯トウモロコシの収量ポテンシャルの動向」 . Crop Science . 39 (6): 1622– 1630. doi : 10.2135/cropsci1999.3961622x . S2CID 39657597. 2009年11月15日時点のオリジナルよりアーカイブ。
- ^マイティ、ラティカンタ (2012).作物植物解剖学. CABI. p. 52. ISBN 978-1-78064-174-4。
- ^ロペス、アデルマリー・プレステス;ノブレガ、ルシア・ヘレン・ペレイラ。パチェコ、ファビオ・パルチェフスキ。クルス=シルバ、クラウディア・タチアナ・アラウホ・ダ(2016)。 「有機栽培下でのベビーコーンの収量と収穫後の品質を考慮したトウモロコシ品種」。バイオサイエンスジャーナル。32 (2): 298–307。土井: 10.14393/BJ-v32n2a2016-26230。
- ^ McCaw, Morgan E; Wallace, Jason G; Albert, Patrice S; Buckler, Edward S; Birchler, James A (2016年9月1日). 「早咲きミニトウモロコシ:種子から種子まで60日で」 . Genetics . 204 (1): 35– 42. doi : 10.1534/genetics.116.191726 . ISSN 1943-2631 . PMC 5012399. PMID 27440866 .
- ^ヴァン・ダインゼ、アレン;サモラ、パブロ。ドゥロー、ピエールマルク。ハイトマン、クリストバル。ジャヤラマン、ディリープクマール。他。 (2018年)。「トウモロコシの在来種における窒素固定は、粘液に関連したジアゾ栄養微生物叢によってサポートされています。 」 PLOS 生物学。16 (8) e2006352。土井:10.1371/journal.pbio.2006352。PMC 6080747。PMID 30086128。
- ^ 「私たちについて」CIMMYT . 2024年3月12日閲覧。
- ^ 「アフリカ向け干ばつ耐性トウモロコシ(DTMA)」 CIMMYT、2022年6月17日。 2024年3月12日閲覧。
- ^ a bチャクラダール、タンミネニ;ヒンドゥー教、ヴェムリ語。レディ、パラコラヌ・スダカール(2017年12月)。「熱帯トウモロコシ改良のためのゲノムベースの育種ツール」(PDF)。ジェネティカ。145 (6): 525–539 .土井: 10.1007/s10709-017-9981-y。PMID 28875394。S2CID 24074330。
- ^ Rashid, Zerka; Singh, Pradeep Kumar; Vemuri, Hindu; Zaidi, Pervez Haider; Prasanna, Boddupalli Maruthi; Nair, Sudha Krishnan (2018年1月10日). 「アジア適応型熱帯トウモロコシにおけるゲノムワイド関連研究により、ソルガムべと病抵抗性に関する新規および探索的ゲノム領域が明らかに」 . Scientific Reports . 8 (1): 366. Bibcode : 2018NatSR...8..366R . doi : 10.1038/ s41598-017-18690-3 . ISSN 2045-2322 . PMC 5762920. PMID 29321632 .
- ^ a b James, Clive (2016). 「バイオテクノロジー/遺伝子組み換え作物の商業化に関する世界の現状:2016年 – ISAAA Brief 52-2016」 . ISAAA. 2017年5月4日時点のオリジナルよりアーカイブ。 2017年8月26日閲覧。
- ^ ISAAA Brief 43-2011 : 概要、2012 年 9 月 9 日取得
- ^ a b c Ostrý, Vladimír; Malíř, František; Pfohl-Leszkowicz, Annie (2015). 「Btトウモロコシと非Bt同質遺伝子トウモロコシのアフラトキシン含有量とヒトおよび動物の健康に関する比較データ – レビュー」Acta Veterinaria Brno . 84 ( 1): 47– 53. doi : 10.2754/avb201585010047 .
- ^ “ISAAA Pocket K No. 2: Plant Products of Biotechnology, 2018” . 2023年1月30日時点のオリジナルよりアーカイブ。 2024年1月9日閲覧。
- ^ポラック、アンドリュー(2000年9月23日)「クラフト、バイオエンジニアリングコーン使用のタコスシェルをリコール」ニューヨーク・タイムズ
- ^フェルナンデス=アルメスト、フェリペ (2011)。世界: 歴史。ロンドン: ペンギン学術。 p. 470.ISBN 978-0-205-75930-9。
- ^ヒル、クリスティーナ・ギッシュ(2020年11月20日)「トウモロコシ、豆、カボチャの『三姉妹』をネイティブアメリカンの農場に戻すことは、人々、土地、そして文化を育む」 The Conversation . 2021年1月9日閲覧。
- ^マン、チャールズ・C.(2011年7月)「綿花(またはアンチョビ)とトウモロコシ」『1491年:コロンブス以前のアメリカ大陸の新発見』(第2版)ニューヨーク:ヴィンテージ・ブックス、 225~229頁、ISBN 978-1-4000-3205-1。
- ^ 「家庭菜園でスイートコーンを育てる」ジョージア大学エクステンション。2024年3月9日閲覧。
- ^スミス、C. ウェイン、ベトラン、ハビエル、ルンゲ、エドワード CA (2004年3月8日). 『トウモロコシ:起源、歴史、技術、生産』ジョン・ワイリー・アンド・サンズ802ページ. ISBN 978-0-471-41184-0。
- ^ George K. Holmes, "Supply and Wages of Farm Labor" Yearbook of the United States Department of Agriculture: 1910 (1911) pp. 190-191 online
- ^ブラウン、ロバート・C. ; ブラウン、トリスタン・R. (2013年12月6日). 『バイオ再生可能資源:農業から新しい製品を生み出す』 . Wiley. PT114. ISBN 978-1-118-52492-3。
- ^ Van Devender, Karl (2011年7月). 「穀物乾燥の概念と選択肢」(PDF) .アーカンソー大学農学部.オリジナル(PDF)から2016年6月1日時点のアーカイブ。 2013年12月15日閲覧。
- ^国際穀物理事会(国際機関)(2013年)「国際穀物理事会市場レポート 2013年11月28日」(PDF)。
- ^ a b「FAOSTAT」。FAO。
- ^ 「2017年のトウモロコシ生産量、作物/地域/生産量(選択リストより)」。国連食糧農業機関統計部(FAOSTAT)。2018年。 2020年3月15日閲覧。
- ^ FAO (2025).世界食料農業統計年鑑2025. FAO. doi : 10.4060/cd4313en . ISBN 978-92-5-140174-3。
- ^ 「トウモロコシの害虫」ユタ州立大学。 2024年1月11日閲覧。
- ^ Mueller, Daren; Pope, Rich編 (2009).トウモロコシ畑ガイド(PDF) .アイオワ州立大学エクステンション. 2024年1月11日時点のオリジナル(PDF)からアーカイブ。 2024年1月11日閲覧。
- ^ 「トウモロコシの病気と障害」マニトバ州農業局。 2024年1月11日閲覧。
- ^ワイズ、キアステン. 「トウモロコシの病気:トウモロコシ北部葉枯れ病」(PDF) .パデュー大学. 2024年1月11日閲覧。
- ^ Juroszek, Peter; von Tiedemann, Andreas (2013). 「気候変動とトウモロコシ病害の将来的な重要性:短評」 . Journal of Plant Diseases and Protection . 120 (2): 49– 56. Bibcode : 2013JPDP..120...49J . doi : 10.1007/BF03356454 . S2CID 87234896 .
- ^ 「米国およびカナダ・オンタリオ州のトウモロコシ病害による被害推定値 — 2022年」 . cropprotectionnetwork.org . 2024年1月11日閲覧。
- ^ホジソン、エリン W. (2008)ユタ州立大学エクステンションおよびユタ州植物害虫診断研究所.ウエスタンコーンルートワーム
- ^ Ostlie, KR; et al. 「Btトウモロコシとヨーロッパアワノメイガ:耐性管理による長期的な成功」ミネソタ大学エクステンションオフィス。2013年9月28日時点のオリジナルよりアーカイブ。
- ^ Reisig, Dominic; Kesheimer, Katelyn; Bateman, Nick; Studebaker, Glenn; Meyer, Ron; et al. (2022年7月22日). 「米国およびカナダ・オンタリオ州におけるトウモロコシ無脊椎動物損失推定値 — 2021」 . doi : 10.31274/cpn-20220722-0 . S2CID 251087338. 2024年1月11日閲覧。
- ^ "fall armyworm, Spodoptera frugiperda (JE Smith)" . entnemdept.ufl.edu . 2017年11月14日閲覧。
- ^ 「PestWeb | Greater Rice Weevil」 . Agspsrv34.agric.wa.gov.au. 2011年9月28日時点のオリジナルよりアーカイブ。 2010年7月29日閲覧。
- ^ Thakur, JN; Rawat, US; Pawar, AD (1987). 「インド・クル(ヒマーチャル・プラデーシュ州)におけるトウモロコシの重篤害虫としてのアワヨトウ( Mythimna separata (Haworth))の初記録と総合的防除に関する提言」.熱帯性害虫管理. 33 (2): 173– 175. doi : 10.1080/09670878709371141 .
- ^ Norton, Don C. (1983年3月). 「トウモロコシの線虫問題」(PDF) .植物病理学. 67 (3): 253– 256. Bibcode : 1983PlDis..67..253N . doi : 10.1094/PD-67-253 .
- ^ Meissle, Michael; Mouron, Patrik; Musa, Tomke; et al. (2010). 「欧州のトウモロコシ生産における害虫、農薬使用、代替策:現状と将来展望」 . Journal of Applied Entomology . 134 (5): 357– 375. doi : 10.1111/j.1439-0418.2009.01491.x . hdl : 11568/710064 . S2CID 73606627 .
- ^ 「コーンスターチ」メリアム・ウェブスター。2016年3月4日時点のオリジナルよりアーカイブ。 2016年5月14日閲覧。
- ^欧州澱粉協会 (2013年6月10日). 「ブドウ糖果糖シロップおよびイソグルコースに関するファクトシート」 . AAF . 2014年6月3日時点のオリジナルよりアーカイブ。 2024年1月11日閲覧。
- ^キニリー、ローラ. 「バーボンの名前の由来とアメリカのネイティブスピリッツに関するヒント」. Smithsonian.com . 2013年6月13日.
- ^トウモロコシ精製協会.コーン油2019年4月12日アーカイブat the Wayback Machine第5版. 2006
- ^ピルチャー、ジェフリー・M. (2012). 「トウモロコシとメキシコの誕生」.プラネット・タコ:メキシコ料理の世界史.オックスフォード大学出版局. p. 27. ISBN 978-0-19-974006-2。
- ^デイビッドソン、アラン (2014). 「メキシコ」.オックスフォード・コンパニオン・トゥ・フード(第3版).オックスフォード大学出版局. pp. 516– 517. ISBN 978-0-19-967733-7。
- ^ピーターソン、ジェームズ (2012). 『野菜 改訂版:300種類以上のレシピを掲載した、最も権威ある購入・準備・調理ガイド』ランダムハウス. p. 184. ISBN 978-1-60774-205-0。
- ^ Nweke, Felix I. 「アフリカにおけるキャッサバの変容」国連食糧農業機関(FAO ) 2024年1月8日閲覧。
- ^ニールセン、L.ミシェル (2007). 『トウモロコシの伝記』 Crabtree Publishing. p. 27. ISBN 978-0-7787-2491-9。
- ^米国食品医薬品局(2024). 「栄養成分表示ラベルおよびサプリメント表示ラベルに記載されている1日あたりの摂取量」 . FDA . 2024年3月27日時点のオリジナルよりアーカイブ。 2024年3月28日閲覧。
- ^ 「表4-7 本報告書で定められたカリウム適正摂取量と2005年DRI報告書で定められたカリウム適正摂取量の比較」 120ページ。Stallings , Virginia A.、Harrison, Meghan、Oria, Maria 編 (2019). 「カリウム:適切な摂取量のための食事摂取基準」.ナトリウムとカリウムの食事摂取基準. pp. 101– 124. doi : 10.17226/25353 . ISBN 978-0-309-48834-1. PMID 30844154 . NCBI NBK545428 .
- ^ “FoodData Central” . fdc.nal.usda.gov . 2019年4月3日時点のオリジナルよりアーカイブ。2021年1月15日閲覧。
- ^ a b「第8章:トウモロコシ食の改善;機関文書『ヒトの栄養におけるトウモロコシ』より」国連食糧農業機関(FAO)1992年。 2017年6月5日閲覧。
- ^ Heuze、V.;トラン、G.エドゥアール、N. Lebas、F. (2017 年 6 月 22 日)。「トウモロコシサイレージ」。 Feedipedia、INRA、CIRAD、AFZ、FAO によるプログラム。
- ^ Heuze、V.;トラン、G.エドゥアール、N. Lebas、F. (2017 年 6 月 21 日)。「トウモロコシの緑の飼料」。 Feedipedia、INRA、CIRAD、AFZ、FAO によるプログラム。
- ^ 「俵型トウモロコシの茎」イリノイ大学アーバナ・シャンペーン校. 2023年12月28日閲覧。
- ^ 「コーンスターチ」(PDF) . コーン精製業者協会. 2013年. 2024年1月9日閲覧。
- ^ Liggett, R. Winston; Koffler, H. (1948年12月). 「微生物学におけるコーンスティープリカー」 . Bacteriological Reviews . 12 (4): 297– 311. doi : 10.1128/MMBR.12.4.297-311.1948 . PMC 180696. PMID 16350125 .
- ^ 「家庭暖房用トウモロコシ:実現しなかったグリーンアイデア」 2015年3月2日。 2015年3月3日時点のオリジナルよりアーカイブ。2017年7月7日閲覧。
- ^ Torres, Andres F.; Slegers, Petronella M.; Noordam-Boot, Cornelie MM; 他 (2016年12月). 「消化率を向上させたトウモロコシ原料は、バイオマスの前処理および糖化のコストと環境への影響を軽減する」 . Biotechnology for Biofuels . 9 (1): 63. Bibcode : 2016BB......9...63T . doi : 10.1186/s13068-016-0479-0 . PMC 4791978. PMID 26981155 .
- ^クレイトン、マーク(2008年1月28日)「世界の食料価格が上昇する中、バイオ燃料が原因か?」クリスチャン・サイエンス・モニター。 2014年10月6日閲覧。
- ^ヘルマン、ホフバウアー;ラインハルト、R.クラウス、ボッシュ。ラインハルト、K.クリスチャン、アイヒャーニヒ (2002 年 1 月)。「バイオマス CHP プラント Güssing - 成功事例」。経済労働省、ニーダーエステルライヒ連邦州およびブルゲンラント連邦州。S2CID 56073239。
- ^ Bassie, Karen (2002). 「Corn Deities and the Complementary Male/Female Principle」. Lowell S. Gustafson、Amelia N. Trevelyan (編). 『古代マヤのジェンダー・アイデンティティと関係』 . Westport, Conn. and London: Bergin&Garvey. pp. 169– 190. 2009年7月10日時点のオリジナルよりアーカイブ。 2007年12月5日閲覧。
- ^ 「トウモロコシの穂軸またはトウモロコシの茎でできた柱と柱頭」。国会議事堂の建築家。 2024年1月11日閲覧。
- ^ 「コーン・パレスの歴史」ミッチェル市。2007年9月29日時点のオリジナルよりアーカイブ。2007年10月15日閲覧。
- ^ Gordon, Ken (2019年9月28日). 「奇妙なものから大切なダブリンの象徴へ、『Field of Corn』が25周年を祝う」 . The Columbus Dispatch . 2019年9月30日時点のオリジナルよりアーカイブ。 2021年12月21日閲覧。
- ^クロアチア国立銀行.クロアチアの硬貨「クーナとリパ」Archived June 22, 2009, at the Wayback Machine : 1 リパ硬貨Archived June 28, 2011, at the Wayback Machine . 2009年3月31日閲覧。
- ^プロエザ、メモリア チレナ。スペイン語で。
- ^ «新しいことはありません» . 2013年。エル・モストラドール。
さらに読む
- バイアリー、デレク. 「ハイブリッドトウモロコシのグローバリゼーション、1921~1970年」『グローバルヒストリージャーナル』 15.1 (2020): 101-122.
- クランプット、シンシア『トウモロコシ:トウモロコシがアメリカ中西部をどう形作ったか』(2015年)
- ボナビア、ドゥッチョ(2013年5月13日)『トウモロコシ:起源、栽培化、そして文化の発展における役割』ケンブリッジ大学出版局、ISBN 978-1-107-02303-1。
外部リンク