海洋食物網

海洋食物網は、海洋生物の食物網です。海洋食物網の基盤には、単細胞藻類や植物プランクトンと呼ばれる植物様生物が存在します。第2栄養段階(一次消費者)は、植物プランクトンを餌とする動物プランクトンが占めています。高次消費者が食物網を完成させます。近年、海洋微生物に関する認識が高まっています。
生息地は食物網の多様性につながります。栄養段階間の相互作用のネットワークは、海洋生態系の機能について多くの情報を提供します。
陸上環境と比較すると、海洋環境のバイオマスピラミッドは底辺が反転しています。特に、消費者(カイアシ類、オキアミ、エビ、餌となる魚類)のバイオマスは、一次生産者のバイオマスよりも大きいです。これは、海洋の一次生産者が急速に成長・繁殖する微小な植物プランクトンであるため、小さな塊でも一次生産速度が速いためです。対照的に、成熟した森林など、多くの重要な陸上一次生産者は成長・繁殖が遅いため、同じ一次生産速度を達成するにははるかに大きな塊が必要になります。この反転により、海洋動物のバイオマスの大部分は動物プランクトンで構成されています。
| 概要シリーズの一部 |
| 海洋生物 |
|---|
 |

食物連鎖と栄養段階
| 海洋食物連鎖(典型) |
|---|
 太陽 太陽 |
| ↓ |
| 植物プランクトン |
| ↓ |
| 草食動物プランクトン |
| ↓ |
| 肉食性動物プランクトン |
| ↓ |
 濾過摂食者 濾過摂食者 |
| ↓ |
 捕食性脊椎動物 捕食性脊椎動物 |
食物網は食物連鎖によって形成されます。海に生息するあらゆる生物は、他の生物の餌となる可能性があります。海における食物連鎖は、通常、太陽エネルギーが植物プランクトンに供給することから始まり、次のような経路を辿ります。
植物プランクトン → 草食性動物プランクトン → 肉食性動物プランクトン →濾過摂食動物→ 捕食性脊椎動物
植物プランクトンは、太陽光をエネルギー源として無機炭素から直接栄養を作り出す能力を持っているため、他の生物を餌として必要としません。このプロセスは光合成と呼ばれ、植物プランクトンは天然に存在する炭素を原形質に変換します。このため、植物プランクトンは海洋食物連鎖の底辺、つまり第一段階における一次生産者と言われています。第一段階にあるため、栄養段階は1(ギリシャ語で「食物」を意味するtrophēに由来)と言われています。植物プランクトンは、食物連鎖の次の栄養段階において、動物プランクトンと呼ばれる微小な動物によって消費されます。
動物プランクトンは食物連鎖の第二栄養段階を構成し、原生動物と呼ばれる微小な単細胞生物、カイアシ類やオキアミなどの小型甲殻類、そして魚類、イカ、ロブスター、カニの幼生などが含まれます。この段階の生物は一次消費者と考えることができます。
一方、小型の草食性動物プランクトンは、大型の捕食性原生動物やオキアミなどの大型の肉食性動物プランクトンや、小型で群れを成し、濾過摂食を行う餌魚類に消費されます。これは食物連鎖の第三栄養段階を構成します。


第4栄養段階は、捕食魚、海棲哺乳類、そして餌となる魚類を捕食する海鳥で構成されています。例としては、メカジキ、アザラシ、カツオドリなどが挙げられます。
アザラシを捕食するシャチや、メカジキを捕食するアオザメなどの頂点捕食者は、第5の栄養段階を構成します。ヒゲクジラは動物プランクトンやオキアミを直接捕食するため、食物連鎖は3~4段階にとどまります。
実際には、栄養段階は通常単純な整数ではありません。なぜなら、同じ消費者種が複数の栄養段階にまたがって餌をとることがよくあるからです。[ 4 ] [ 5 ]例えば、大型の海洋脊椎動物は小型の捕食魚を食べることもありますが、濾過摂食動物も食べることがあります。アカエイは甲殻類を食べますが、アオシュモクザメは甲殻類とアカエイの両方を食べます。動物は互いに食べ合うこともあります。タラは小型のタラとザリガニを食べ、ザリガニはタラの幼生を食べます。幼生の食性、ひいては栄養段階は、成長するにつれて変化することがあります。
水産学者ダニエル・ポーリーは、一次生産者とデトリタスを栄養段階1 、草食動物とデトリタス食動物(一次消費者)を栄養段階2、二次消費者を栄養段階3、といった具合に設定している。あらゆる消費者種における栄養段階TLの定義は[ 6 ]である。

ここで、 は獲物jの栄養段階の割合であり、 はiの食事におけるjの割合を表す。海洋生態系の場合、ほとんどの魚類やその他の海洋消費者の栄養段階は2.0から5.0の間の値をとる。上限値5.0は、大型魚類であっても珍しいが[ 7 ]、ホッキョクグマやシャチなどの海洋哺乳類の頂点捕食者では見られる。[ 8 ]対照的に、人間の平均栄養段階は約2.21で、豚やカタクチイワシとほぼ同じである。[ 9 ] [ 10 ]


分類別
一次生産者

海洋食物網の基盤には、植物プランクトンと呼ばれる単細胞藻類やその他の植物様生物が存在します。植物プランクトンは、形態、大きさ、色素の種類に基づいて多様な分類群に分類される微小な独立栄養生物のグループです。海洋植物プランクトンは主に光合成独立栄養生物として太陽光が当たる表層水に生息し、炭素を固定して酸素を生成するために窒素やリンなどの栄養素、そして太陽光を必要とします。しかし、一部の海洋植物プランクトンは深海、特に深海噴出孔付近に生息し、硫化水素、第一鉄、アンモニアなどの無機電子源を利用する化学合成独立栄養生物として生息します。[ 12 ]
生態系を理解するには、その食物網が物質とエネルギーの流れをどのように決定するかを知らなければなりません。植物プランクトンは、無機化合物を有機化合物に変換することで独立栄養的にバイオマスを生産します。このように、植物プランクトンは海洋の他のすべての生物を支えることで、海洋食物網の基盤として機能します。海洋食物網における2番目の中心的なプロセスは、微生物ループです。このループは海洋細菌と古細菌を分解し、有機物と無機物を再ミネラル化し、その後、その産物を外洋性食物網内で、または海底に海洋堆積物として堆積させることでリサイクルします。 [ 13 ]
海洋植物プランクトンは海洋食物網の基盤を形成し、光合成による世界の炭素固定と酸素生産の約半分を占め[ 14 ]、地球全体の炭素循環において重要な役割を果たしています。[ 15 ]陸上の植物と同様に、植物プランクトンはクロロフィルなどの集光性色素を用いて光合成を行い、大気中の二酸化炭素を吸収して燃料となる糖を生成します。水中のクロロフィルは、水が太陽光を反射・吸収する方法を変化させるため、科学者は植物プランクトンの量と位置をマッピングすることができます。これらの測定値は、科学者に海洋環境の健全性に関する貴重な知見を提供し、海洋炭素循環の研究にも役立ちます。[ 11 ]
植物プランクトンは、食べられる前に死んだ場合、マリンスノーの一部として有光層を下り、深海に沈みます。このように、植物プランクトンは毎年約20億トンの二酸化炭素を海に吸収・固定しており、海は吸収された炭素の約90%を吸収する二酸化炭素吸収源となっています。[ 16 ]海は世界の酸素の約半分を生成し、大気中の50倍の二酸化炭素を吸収しています。[ 17 ]

植物プランクトンの中には、シアノバクテリア門に属する細菌が存在します。海洋シアノバクテリアには、既知の光合成生物の中で最も小さいものが含まれます。その中で最も小さいのがプロクロロコッカスで、体長はわずか0.5~0.8マイクロメートルです。[ 18 ]個体数で言えば、プロクロロコッカスは地球上で最も数が多い種であると考えられます。表層海水1ミリリットルには10万個以上の細胞が含まれることがあります。世界中に推定で数千億(10の27乗)の個体が存在するとされています。[ 19 ]プロクロロコッカスは北緯40度から南緯40度の間に広く分布し、海洋の貧栄養地域(栄養塩の少ない地域)で優占しています。[ 20 ]この細菌は地球の大気中の酸素の約20%を占めています。[ 21 ]
- 植物プランクトンは海洋食物連鎖の基盤を形成する
 植物プランクトン
植物プランクトン- 渦鞭毛藻類
- 珪藻類



海洋では、一次生産の大部分は藻類によって行われています。これは、一次生産の大部分が維管束植物によって行われている陸上とは対照的です。藻類は、単独の浮遊細胞から付着性海藻まで多岐にわたりますが、維管束植物は、海草やマングローブなどのグループに代表されます。海草や海藻などの大規模な生産者は、主に沿岸域や浅瀬に限定され、そこでは基質に付着し、光合成層内に留まります。しかし、藻類による一次生産の大部分は、植物プランクトンによって行われています。
このように、海洋環境において、第一階層の栄養段階は主に植物プランクトンによって占められています。植物プランクトンとは、海中を漂う微小な漂流生物で、主に単細胞藻類です。ほとんどの植物プランクトンは肉眼では1つ1つ見ることができません。数が多いと、水が(しばしば緑色に)変色して見えることがあります。植物プランクトンは主に光合成によってバイオマスを増殖させるため、太陽光が当たる海面(有光層)に生息します。
植物プランクトンの中で最も重要なグループには、珪藻類と渦鞭毛藻類が含まれます。珪藻類は特に海洋において重要であり、ある推定によると、海洋の一次生産量全体の最大45%を占めています。[ 22 ]珪藻類は通常は顕微鏡でしか見えませんが、種によっては体長2ミリメートルに達するものもあります。
一次消費者

第 2 栄養段階 (一次消費者) は、植物プランクトンを食べる動物プランクトンが占めています。動物プランクトンと植物プランクトンが食物連鎖の底辺を成し、世界の主要漁場のほとんどを支えています。動物プランクトンは、海洋表層水で植物プランクトンと一緒に見られる小さな動物が多く、小さな甲殻類や、魚の仔魚や稚魚(孵化したばかりの魚)が含まれます。ほとんどの動物プランクトンは濾過摂食者で、付属肢を使って水中の植物プランクトンを濾過します。大型の動物プランクトンの中には、小型の動物プランクトンを食べるものもあります。捕食者を避けるためにジャンプできる動物プランクトンもいますが、実際には泳ぐことはできません。植物プランクトンと同様に、動物プランクトンも海流や潮流、風に乗って漂います。動物プランクトンは繁殖力が強く、好条件下では1日に最大30%まで個体数が増加することがあります。多くの動物プランクトンは短命ながらも生産的で、急速に成熟します。
寡毛類は、襟と折り返しのような形状の顕著な口繊毛を持つ繊毛虫の一種です。海洋プランクトン群集には非常に多く見られ、通常は1mlあたり約1匹の濃度で存在します。彼らは海で最も重要な草食動物であり、食物連鎖の第一段階を担っています。[ 23 ]
動物プランクトンの中で特に重要なグループには、カイアシ類とオキアミがあります。カイアシ類は、海洋および淡水域に生息する小型甲殻類です。海における最大のタンパク質源であり[ 24 ]、餌魚の重要な餌となります。オキアミは次に大きなタンパク質源です。オキアミは特に大型の捕食性動物プランクトンで、より小型の動物プランクトンを餌とします。つまり、オキアミは餌魚とともに、実際には第3栄養段階、つまり二次消費者に属します。
- 動物プランクトンは海洋食物連鎖の第二段階を形成する
- 節足動物
- 小さなエビのような甲殻類
- プランクトン性の幼生イカ




海中のプランクトンの大部分は、植物プランクトンと動物プランクトンで構成されています。プランクトンとは、海を漂う小さな漂流生物全般を指します(ギリシャ語で「planktos」は「放浪者」または「漂流者」を意味します)。定義上、プランクトンに分類される生物は海流に逆らって泳ぐことができず、周囲の流れに抵抗して自分の位置を制御することができません。海洋環境において、最初の2つの栄養段階は主にプランクトンによって占められています。プランクトンは生産者と消費者に分けられます。生産者は植物プランクトン(ギリシャ語で「phyton」は「植物」)であり、植物プランクトンを食べる消費者は動物プランクトン(ギリシャ語で「zoon」は「動物」)です。
クラゲは泳ぎが遅く、ほとんどの種がプランクトンの一部を形成する。伝統的に、クラゲは栄養段階の行き止まりであると考えられてきた。主に水をベースとした体制であるため、海洋生態系への影響は限定的であると一般的に考えられており、マンボウやオサガメなどの特殊な捕食者の注目を集めていた。[ 26 ] [ 25 ]この見解は最近異論が出ている。クラゲ、およびより一般的にはサルパや有櫛動物を含むゼラチン状の動物プランクトンは、非常に多様で、壊れやすく硬い部分がなく、観察や監視が難しく、急速な個体数の変動があり、多くの場合、海岸から遠く離れた場所や海の深いところに生息している。科学者が捕食者の腸内でクラゲを検出して分析することは困難である。なぜなら、クラゲは食べられるとドロドロになり、急速に消化されるからである。[ 26 ]しかし、クラゲは大量に発生し、マグロ、スピアフィッシュ、メカジキなどの魚類や、タコ、ナマコ、カニ、端脚類などの無脊椎動物の餌として主要な役割を果たしていることが示されています。[ 27 ] [ 25 ] 「クラゲはエネルギー密度が低いにもかかわらず、消化が速く、捕獲コストが低く、入手しやすく、エネルギーに富んだ成分を選択的に摂取するため、捕食者のエネルギー収支へのクラゲの貢献は想定よりもはるかに大きい可能性があります。クラゲを餌とすることで、海洋捕食者はプラスチックを摂取しやすくなる可能性があります。」[ 25 ]
高次の消費者

- 海洋無脊椎動物
- 魚
- 餌魚類:餌魚類は海洋食物網において中心的な位置を占める。餌魚類が捕食する生物は低栄養段階にあり、餌魚類を捕食する生物は高栄養段階にある。餌魚類は食物網の中位に位置し、高栄養段階の魚類、海鳥、哺乳類の主要な捕食対象となっている。[ 28 ]
- 捕食魚
- すり身魚
- その他の海洋脊椎動物
2010年、研究者たちはクジラが「ホエールポンプ」と呼ばれるプロセスを使って、海の深層から栄養分を表層へと運ぶことを発見しました。[ 29 ]クジラはオキアミが生息する深層で餌を食べますが、呼吸のために定期的に表層に戻ります。そこでクジラは窒素と鉄分を豊富に含む液体を排泄します。排泄された液体は沈むことなく表層に留まり、植物プランクトンがそれを消費します。メイン湾では、ホエールポンプが河川よりも多くの窒素を供給しています。[ 30 ]

,_Belmont_-_geograph.org.uk_-_529175.jpg/1280px-Gannet_(Morus_bassana),_Belmont_-_geograph.org.uk_-_529175.jpg)
- クジラポンプ栄養循環

微生物

近年、海洋微生物が海洋生態系において、これまで考えられていたよりもはるかに大きな役割を果たしていることが認識されつつあります。メタゲノミクスの発展により、研究者はこれまで隠されていた微生物の多様性を明らかにすることが可能になり、微生物の世界を観察するための強力なレンズが提供されるとともに、生物界への理解に革命をもたらす可能性を秘めています。[ 32 ]メタバーコーディングによる食物網分析技術は、食物網をより高い分類学的解像度で再構築するために用いられており、相互作用網のより深い複雑さを明らかにしています。[ 33 ]
微生物は海洋食物網において重要な役割を果たしている。ウイルスシャント経路は、海洋微生物の粒子状有機物(POM)を微生物が容易に吸収できる溶存有機物(DOM)にリサイクルすることで、栄養段階の上昇を防ぐメカニズムである。 [ 34 ]ウイルスシャントは、単一種の海洋微生物が微小環境を支配するのを防ぐことで、微生物生態系の多様性を維持するのに役立つ。[ 35 ]ウイルスシャント経路によってリサイクルされるDOMは、他の主要な海洋DOM発生源によって生成される量に匹敵する。[ 36 ]
一般的に、溶存有機炭素(DOC)は、細菌による溶解、植物プランクトンからの固定炭素の漏出または滲出(例:珪藻類の粘液性細胞外ポリマー)、突然の細胞老化、動物プランクトンの乱獲、水生動物による老廃物の排泄、または陸生植物および土壌からの有機粒子の分解または溶解によって海洋環境に導入される。[ 37 ]微生物ループ内の細菌は、この粒子状のデトリタスを分解し、このエネルギーに富む物質を成長に利用する。海洋生態系の有機物の95%以上はポリマーの高分子量(HMW)化合物(例:タンパク質、多糖類、脂質)で構成されているため、高栄養段階のほとんどの海洋生物が容易に利用できるのは、総溶存有機炭素のごく一部である。これは、溶存有機炭素がほとんどの海洋生物に直接利用できないことを意味する。海洋細菌はこの有機炭素を食物網に導入し、その結果、より高い栄養段階で追加のエネルギーが利用できるようになります。




.jpg/1280px-CroV_TEM_(cropped).jpg)


- ウイルス
ウイルスは「地球上で最も豊富な生物学的実体」であり、[ 41 ]、特に地球の表面積の70%以上を占める海洋においてその存在が顕著である。[ 41 ] [ 42 ] 1989年に海水1ミリリットル中に約100個の海洋ウイルスが存在することが明らかになり、 [ 43 ]海洋環境におけるウイルスの多様性と役割を理解するきっかけとなった。[ 42 ]現在、ウイルスは微生物群集の動態、宿主の代謝状態、宿主の溶解を介した生物地球化学的循環を制御することで、海洋生態系において重要な役割を果たしていると考えられている。[ 41 ] [ 42 ] [ 44 ] [ 45 ]
巨大海洋ウイルスCroV が海洋動物鞭毛藻Cafeteria roenbergensisに感染し、溶解により死滅させる。[ 46 ] Cafeteria roenbergensis は水中の細菌を餌とするため、沿岸生態系に影響を及ぼしている。CroVの広範な感染によりCafeteria roenbergensisの数が少なくなると、細菌の個体数が飛躍的に増加する。 [ 47 ] CroVがC. roenbergensisの自然個体群に与える影響は不明であるが、このウイルスは宿主特異性が非常に高く、近縁の他の生物には感染しないことが判明している。[ 48 ] Cafeteria roenbergensis は、第 2 のウイルスであるMavirus のビロファージにも感染する。これはサテライトウイルスであり、別の特定のウイルス、この場合は CroV が存在する場合にのみ複製することができる。[ 49 ]このウイルスはCroVの複製を阻害し、 C. roenbergensis細胞の生存につながります。MavirusはC. roenbergensis細胞のゲノムに統合することができ、それによって集団に免疫を与えます。[ 50 ]
菌類
寄生性ツボカビは、マイコループと呼ばれるプロセスによって、大型の非食用植物プランクトンから動物プランクトンへ物質を移すことができます。ツボカビの遊走子は、その大きさ(直径2~5μm)、形状、栄養価(多価不飽和脂肪酸とコレステロールが豊富)の点で、動物プランクトンにとって優れた餌となります。宿主植物プランクトンの大きなコロニーも、ツボカビの感染によって断片化され、動物プランクトンにとって食用となることがあります。[ 51 ]
- 菌ループ(菌類ループ)の図


寄生菌類は腐生菌類と同様に、植物プランクトンの有機炭素を直接同化します。遊走子を放出することで、菌類は動物プランクトンとの栄養連鎖(菌類ループ)を橋渡しします。粒子状および溶存有機炭素を変化させることで、菌類は細菌や微生物ループに影響を及ぼすことができます。これらのプロセスは、マリンスノーの化学組成を変化させ、ひいては生物炭素ポンプの機能に影響を与える可能性があります。[ 52 ] [ 53 ]
生息地別
外洋性網

外洋性生態系については、ルジャンドルとラスールザダガンが1995年に、草食動物の食物連鎖と微生物ループを食物網の末端メンバーとする連続した栄養経路を提唱した。[ 55 ]古典的な線形食物連鎖の末端メンバーは、動物プランクトンがより大きな植物プランクトンを摂食し、それに続いてより大きな動物プランクトンまたは他の捕食者が動物プランクトンを捕食することを伴う。このような線形食物連鎖では、捕食者は植物プランクトンバイオマスの増加(植物プランクトン、草食動物、捕食者のシステムの場合)または植物プランクトンのバイオマスの減少(4つのレベルのシステムの場合)につながる可能性がある。したがって、捕食者の豊富さの変化は、栄養カスケードにつながる可能性がある。[ 56 ]微生物ループの末端メンバーには、基本資源としての植物プランクトンだけでなく、溶存有機炭素も含まれる。[ 57 ]溶存有機炭素は従属栄養細菌の成長に利用され、より大きな動物プランクトンに捕食されます。その結果、溶存有機炭素は細菌-微小動物プランクトンループを経て動物プランクトンに変換されます。これらの2つの末端炭素処理経路は複数のレベルでつながっており、小型植物プランクトンは微小動物プランクトンに直接消費されます。[ 54 ]
右の図に示されているように、溶存有機炭素は、有機炭素の一次生産者と消費者の両方によって、さまざまな生物によって複数の方法で生成されます。一次生産者による DOC の放出は、漏出によって受動的に、また栄養塩制限中の不均衡な成長中に能動的に発生します。[ 58 ] [ 59 ]植物プランクトンから溶存有機炭素プールへの別の直接的な経路は、ウイルスの溶解です。[ 60 ]海洋ウイルスは、特に温暖な低緯度の海域で、海洋植物プランクトンの死亡の主な原因です。草食動物によるずさんな摂食と消費者による獲物の不完全な消化も、溶存有機炭素の他の発生源です。従属栄養微生物は、細胞外酵素を使用して粒子状有機炭素を可溶化し、これおよびその他の溶存有機炭素資源を成長と維持に使用します。微生物の従属栄養生産の一部は微小動物プランクトンによって使用されます。従属栄養コミュニティの別の部分は激しいウイルス溶解にさらされ、これが再び溶存有機炭素の放出を引き起こします。微生物ループの効率は複数の要因に依存しますが、特に従属栄養微生物の死亡率に対する捕食とウイルス溶解の相対的な重要性に依存します。[ 54 ]
- 外洋性食物網
![海洋食物網と生物ポンプ。海洋の生物ポンプと海洋食物網のつながり、そして船舶、衛星、自律走行車からこれらの構成要素を遠隔的に採取する能力。水色の海水は有光層、濃い青色の海水は薄明層を表す。[61]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/13/Export_Processes_in_the_Ocean_from_Remote_Sensing.jpg/1280px-Export_Processes_in_the_Ocean_from_Remote_Sensing.jpg)
- 中深海食物網
- 中深海魚種が地球の炭素収支に与える影響。[ 62 ] DVM = 日周垂直移動、NM = 非移動。
![中深海生物種が地球の炭素収支に与える影響[62] DVM = 日周垂直移動、NM = 非移動。](/wiki/File:Mesopelagic_species_impact_on_global_carbon_budget.png)
![中深海性イシモチ類は地球上で最も豊富な脊椎動物である可能性があるが、それらについてはほとんど知られていない。[63]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/ac/Sigmops_bathyphilus1.jpg/1280px-Sigmops_bathyphilus1.jpg)


科学者たちは、水深200メートルから1,000メートルの中深層という、これまでほとんど知られていなかった薄明帯の詳細な調査を始めています。この層は、毎年約40億トンの二酸化炭素を大気から除去する役割を果たしています。中深層には、海洋魚類のバイオマスの大部分が生息しています。[ 63 ]
2017年の研究によると、中層魚類の中で最も多様な獲物を捕食しているのはクラゲ類で、次いでフィソネクト・シーフォノフォア、クテノフォア、頭足類が続く。いわゆる「ゼリー網」の重要性はようやく理解され始めたばかりだが、クラゲ類、クテノフォア、クテノフォアは、捕食魚類やイカ類と同様の生態学的影響を及ぼす深層食物網の重要な捕食者となり得るようだ。従来、ゼラチン質の捕食者は海洋栄養経路の供給者としては無力だと考えられてきたが、深層食物網において重要な不可欠な役割を果たしているようだ。[ 64 ]重要な能動輸送機構である日周垂直移動により、中層動物プランクトンは大気から二酸化炭素を隔離するだけでなく、他の中層魚類の炭素需要も供給することができる。[ 65 ]
2020年の研究では、温室効果ガスの排出量が削減されたとしても、2050年までに深海における地球温暖化の進行速度は現在の7倍になる可能性があると報告されています。中層および深層での温暖化は、海洋生物が生存可能な温度を維持するために移動する必要があるため、深海の食物網に重大な影響を及ぼす可能性があります。[ 66 ] [ 67 ]
- 夕暮れ時の魚が海洋生態系に新たな光を当てるThe Conversation、2014年2月10日。
- 数兆単位の海洋ミステリーニューヨーク・タイムズ、2015 年 6 月 29 日。
- 中深海魚 - 2010年のマラスピナ周航遠征。[ 68 ] [ 69 ]

海面では

海洋表層生息地は、海洋と大気の界面に位置する。海洋表層のバイオフィルム状の生息地には、一般にニューストンと呼ばれる表層生息性微生物が生息する。この広大な気水界面は、地球の表面積の70%以上に及ぶ主要な気水交換過程が交わる地点に位置する。海洋表層ミクロ層の細菌、いわゆるバクテリオニューストンは、温室効果ガスの気海間ガス交換、気候活性海洋エアロゾルの生成、海洋のリモートセンシングなどの実用化により注目されている。[ 71 ]特に興味深いのは、微生物の生化学的プロセスによる界面活性剤(界面活性物質)の生成と分解である。外洋における界面活性剤の主な発生源には、植物プランクトン、[ 72 ]陸地からの流出水、大気からの沈着物などがある。[ 71 ]
有色の藻類ブルームとは異なり、界面活性剤関連細菌は海洋カラー画像では見えない場合があります。合成開口レーダーを用いてこれらの「目に見えない」界面活性剤関連細菌を検出できることは、雲、霧、日光など、あらゆる天候条件において計り知れない利点があります。[ 71 ]これは特に強風時に重要です。なぜなら、このような状況では、大気と海洋間のガス交換と海洋エアロゾル生成が最も激しくなるからです。したがって、カラー衛星画像に加えて、SAR衛星画像は、海洋と大気の境界における生物物理学的プロセス、大気と海洋間の温室効果ガス交換、そして気候に影響を与える海洋エアロゾル生成の全体像について、さらなる知見を提供してくれる可能性があります。[ 71 ]
海底で

海底(底生)の生息地は、海洋と地球内部の境界面にあります。
- 浸透と通気

沿岸ウェブ
 |
沿岸水域には、河口域と大陸棚上の水域が含まれます。これらは全海洋面積の約8%を占め[ 77 ]、海洋生産力の約半分を占めています。富栄養化を決定づける主要な栄養素は、沿岸水域の窒素と湖沼のリンです。どちらもグアノ(海鳥の糞)に高濃度で含まれており、グアノは周囲の海や隣接する湖の肥料として作用します。尿酸は主要な窒素化合物であり、その鉱化作用によって様々な形態の窒素が生成されます[ 78 ] 。
生態系は、一見すると明確な境界があるように見えても、隣接する他のシステムから独立して機能することはめったにありません。[ 79 ]生態学者は、生態系間のエネルギーと栄養素の輸送が植物や動物の個体群や群集に重要な影響を及ぼすことをますます認識しています。[ 80 ] [ 81 ]そのよく知られた例は、海鳥が繁殖島で海洋由来の栄養素を糞便(グアノ)の形で濃縮する方法です。これには約15~20%の窒素(N)と10%のリンが含まれています。[ 82 ] [ 83 ] [ 84 ]これらの栄養素は陸上生態系の機能と動態を劇的に変え、一次生産性と二次生産性の向上を支えることができます。[ 85 ] [ 86 ]しかし、多くの研究で様々な分類群にわたってグアノ堆積による陸生成分の窒素濃縮が実証されているものの、[ 85 ] [ 87 ] [ 88 ] [ 89 ]海洋生態系への逆作用を研究した研究はごくわずかであり、これらの研究のほとんどは温帯地域と高栄養水域に限定されていました。[ 82 ] [ 90 ] [ 91 ] [ 92 ]熱帯地方では、サンゴ礁は繁殖する海鳥の個体数が多い島に隣接して見られ、海鳥由来の栄養素が周辺海域に輸送されることで、局所的な栄養濃縮の影響を受ける可能性があります。熱帯海洋生態系に対するグアノの影響に関する研究では、グアノ由来の窒素が海水とサンゴ礁の一次生産者を豊かにすることが示唆されています。[ 90 ] [ 93 ] [ 94 ]
造礁サンゴは窒素を必須要件とし、窒素が一次生産性の主要な制限栄養素である栄養分の乏しい熱帯海域[ 95 ]で繁栄しているため、 [ 96 ]サンゴはこの元素を節約するための特別な適応を発達させてきました。造礁サンゴの定着と維持は、周囲の海から溶存無機窒素(アンモニウムと硝酸塩)を吸収して保持できる単細胞渦鞭毛藻類、 Symbiodinium spp.(褐虫藻)との共生に一部依存しています。 [ 97 ] [ 98 ] [ 99 ]これらの褐虫藻は動物の排泄物をリサイクルし、その後、アミノ酸、[ 100 ]アンモニウムまたは尿素としてサンゴの宿主に返します。[ 101 ]サンゴは窒素に富んだ堆積粒子[ 102 ] [ 103 ]やプランクトンも摂取できます。 [ 104 ] [ 105 ]沿岸の富栄養化と過剰な栄養供給はサンゴに強い影響を与え、骨格の成長を低下させる可能性がある。[ 98 ] [ 106 ] [ 107 ] [ 108 ] [ 94 ]


上図の右側では、(1) アンモニア化によりNH 3とNH 4 +が生成され、(2) 硝化によりNH 4 +の酸化によりNO 3 −が生成されます。(3) 海鳥の糞に典型的なアルカリ性条件下では、NH 3は急速に揮発してNH 4 +に変換され、(4) コロニー外に輸送され、湿性沈着によって遠方の生態系に輸出され、そこで富栄養化が起こります。リン循環はより単純で、移動性は低下しています。この元素は海鳥の糞中にさまざまな化学形で存在しますが、最も移動性が高く、生物学的に利用可能なのはオルトリン酸であり、(5)地下水または表層水によって浸出する可能性があります。[ 78 ]








DNAバーコーディングは、網のノードにおける分類学的解像度を向上させた食物網構造を構築するために用いることができます。これにより、より具体的な種の同定が可能になり、誰が誰を食べるのかをより明確に把握できるようになります。「DNAバーコードとDNA情報は、より大規模な相互作用網の構築に向けた新たなアプローチを可能にし、適切なサンプルサイズを達成するためのいくつかの障害を克服する可能性があります。」[ 33 ]
種の同定に新たに応用された方法はDNAメタバーコーディングである。形態による種の同定は比較的困難であり、多くの時間と専門知識を必要とする。[ 118 ] [ 119 ]ハイスループットシーケンシングDNAメタバーコーディングは、以前のDNA増幅で選択されたグループ特異的プライマーに関して、サンプル全体の分類学的割り当てと同定を可能にする。
- 微生物DNAバーコーディング
- 藻類DNAバーコーディング
- 魚類DNAバーコーディング
- 食事評価におけるDNAバーコーディング
- ケルプの森
- バーンズ, ジャレット E.; レイノルズ, パメラ L.; スタコヴィッツ, ジョン J. (2007). 「侵略と絶滅が沿岸海洋食物網を再構築する」. PLOS ONE . 2 (3): e295. Bibcode : 2007PLoSO...2..295B . doi : 10.1371/journal.pone.0000295 . PMC 1808429. PMID 17356703 .
極地のウェブ


北極と南極の海洋システムは地形構造が大きく異なり、その結果、食物網構造も大きく異なります。[ 120 ]北極と南極の表層性食物網はどちらも、少数の主要種によって大きく制御される特徴的なエネルギーフローを持っています。しかし、どちらにも単一の包括的な食物網は存在しません。代替経路は、回復力とエネルギーフローの維持に重要です。しかし、これらのより複雑な代替経路は、上位栄養段階の種へのエネルギーフローを少なくします。「食物網の構造は地域によって類似しているかもしれませんが、中位栄養段階を支配する個々の種は極域によって異なります。」[ 121 ]



- 北極


北極の食物網は複雑です。海氷の減少は、藻類やプランクトンから魚類、哺乳類に至るまで、食物網全体に影響を及ぼす可能性があります。気候変動が特定の種に与える影響は、食物網全体に波及し、他の幅広い生物にも影響を及ぼす可能性があります。…海氷の減少は、ホッキョクグマの主要な生息地の面積を縮小することで個体群に悪影響を及ぼすだけでなく、食物網効果を通じても悪影響を及ぼしています。北極の海氷期間と面積の減少は、氷の中の栄養豊富なポケットで繁殖するアイスアルジーの減少につながります。これらのアイスアルジーは動物プランクトンに食べられ、動物プランクトンはさらにホッキョクタラに食べられます。ホッキョクタラは、アザラシを含む多くの海洋哺乳類にとって重要な食料源です。アザラシはホッキョクグマの餌となります。したがって、アイスアルジーの減少はホッキョクグマの個体群減少に寄与する可能性があります。[ 122 ]
2020年、研究者らは、過去20年間の北極海における一次生産の測定結果から、植物プランクトンの濃度上昇により、約60%の増加が示されたと報告しました。研究者らは、他の海域から新たな栄養素が流入しているという仮説を立て、これは北極海が将来、より高次の栄養段階の生産とさらなる炭素固定を支えられる可能性があることを意味していると示唆しています。 [ 123 ] [ 124 ]





- 南極
- 氷の下の南極クラゲDiplulmaris antarctica
- ロス海の重要な植物プランクトンである藻類Phaeocystis antarcticaのコロニー。海氷が後退した後の早い時期に藻類が優勢となり、大量の炭素を排出する。[ 127 ]
.jpg/1280px-Ice_planet_and_antarctic_jellyfish_(crop).jpg)
![ロス海の重要な植物プランクトンである藻類ファエオシスティス・アンタルクティカのコロニー。海氷が後退した後の早い時期に藻類が優勢となり、大量の炭素を排出する。[127]](/wiki/File:Phaeocystis.png)
![南極周極海流全域に生息する羽状珪藻類Fragilariopsis kerguelensisは、地球規模の珪酸塩ポンプの重要な駆動力となっている。[128]](http://upload.wikimedia.org/wikipedia/commons/thumb/4/4f/Fragilariopsis_kerguelensis.jpg/1280px-Fragilariopsis_kerguelensis.jpg)





極性微生物
多様な地形に加え、極寒の気候にもかかわらず、極地の水域は微生物が豊富に生息しています。氷河下の地域でさえ、細胞生命はこれらの極限環境に適応しており、地球上の初期の微生物の痕跡が残っている可能性があります。これらの極地のほとんどでは大型動物の摂食が限られているため、ウイルスは重要な死亡要因として認識されつつあり、それによって栄養素の生物地球化学的循環に影響を与え、ひいては季節的および空間的なスケールで生物群集の動態に影響を与えています。[ 45 ]
微生物は北極と南極の食物網の中心です。これらの極地環境には、細菌、古細菌、真核生物など、多様な微生物群集が存在し、ウイルスとともに極地生態系の重要な構成要素となっています。[ 134 ] [ 135 ] [ 136 ]これらは氷河下湖やクリオコナイトの穴など、様々な生息地で見られ、これらの極地の寒冷なバイオームは代謝的に多様な微生物で満ち溢れ、活発な生物地球化学的循環の場となっています。[ 137 ] [ 138 ] [ 139 ]地球表面の約5分の1を占め、人間にとって住みにくいこれらの環境は、独特の微生物群集の生息地となっています。[ 134 ] [ 139 ] [ 140 ]両地域の常在微生物相の類似性はわずか 30% 程度だが、極地の海洋の連結性が限られていることや、淡水供給(それぞれ南極海と北極海に流れ込む氷河融解物と河川に由来)の違いを考えれば、必ずしも驚くことではない。 [ 140 ]分離は距離だけによるものではない。南極は強い南極環流によって駆動される南極海に囲まれているのに対し、北極は陸地に囲まれている。このような異なる地形は、約 4000 万~2500 万年前に 2 つの大陸が地球の反対の極地に移動したことで生じた。磁気および重力データは、 1 億 4500 万年前から 1 億 6100 万年前にアメラシア盆地とユーラシア盆地の影響を受けて北極が陸地に囲まれた冷たい極地へと進化したことを示している。[ 141 ] [ 142 ] [ 143 ]南極大陸は、南極海に囲まれた陸塊である超大陸ゴンドワナの分裂によって形成されました。 [ 134 ] [ 144 ]南極大陸は永久に氷河に覆われており、その面積のわずか0.4%が湖や池が点在する露出した陸地で構成されています。[ 45 ]
これらの環境に存在する微生物(原核生物と真核生物の両方)は、両極間で大きく異なります。[ 140 ] [ 145 ]例えば、南極海の表層水群集の細菌操作分類単位(OTU)の78%と北極海の70%は、各極に固有です。[ 140 ]極域は時間的にも空間的にも変化に富んでいます。小サブユニット(SSU)rRNA遺伝子のV6領域の解析により、北極と南極海の44の極地サンプルから約40万の遺伝子配列と11,000を超えるOTUが得られました。これらのOTUは、両極域で別々にクラスターを形成しており、さらに、異なる環境(沿岸と外洋)と異なる季節の極地細菌プランクトン群集だけでも大きな違いが見られます。 [ 140 ] [ 45 ]
極地は断絶した食物網を特徴としており、生態系機能におけるウイルスの役割は、海洋食物網の他の地域よりもさらに大きいと考えられます。ウイルスの多様性については依然として比較的研究が進んでおらず、極地の生物群集にどのような影響を与えるかは十分に理解されていません。[ 138 ]特に栄養循環への影響は顕著です。[ 136 ] [ 146 ] [ 147 ] [ 45 ]
基礎種とキーストーン種
.jpg/1280px-Kelp_Forest_(12801115735).jpg)

.jpg/1280px-California_Mussels_(2513978279).jpg)
 ヒトデが現代の生態系に与えた影響– PBSのネイチャー
ヒトデが現代の生態系に与えた影響– PBSのネイチャー 基礎種の概念は1972年にポール・K・デイトン[ 149 ]によって提唱され、彼はそれを海洋無脊椎動物および藻類群集の特定のメンバーに適用しました。いくつかの場所での研究から、海洋群集の他のメンバーに不均衡な影響を与える活動を持つ少数の種が存在し、したがってそれらの種が群集の回復力の鍵となることが明らかになりました。デイトンの見解は、基礎種に焦点を当てることで、群集全体の反応(汚染など)を同時に追跡するという極めて困難な作業を行う代わりに、より簡略化されたアプローチで、群集全体が汚染などの撹乱にどのように反応するかをより迅速に理解できるというものでした。
基盤種とは、生態系群集を構築し、その環境を形作り、生態系を定義する上で支配的な役割を果たす種である。このような生態系は、海草藻場、カキ場、サンゴ礁、ケルプ林、マングローブ林など、基盤種にちなんで名付けられることが多い。 [ 150 ]例えば、アカマングローブはマングローブ林によく見られる基盤種である。マングローブの根は、フエダイなどの稚魚の生育場所となっている。[ 151 ]基盤種は食物網のどの栄養段階にも属することができるが、生産者である傾向がある。[ 152 ]

 ラッコ対ウニ–ピュー研究所
ラッコ対ウニ–ピュー研究所 キーストーン種の概念は、1969年に動物学者ロバート・T・ペインによって提唱されました。[ 153 ] [ 154 ]ペインはこの概念を、ヒトデやムール貝などを含む潮間帯(満潮線と干潮線の間)の海洋無脊椎動物間の関係性に関する自身の観察と実験を説明するために提唱しました。ヒトデの中には、他に天敵がいないウニ、ムール貝、その他の貝類を捕食するものがあります。ヒトデが生態系から排除されると、ムール貝の個体数が制御不能に爆発的に増加し、他のほとんどの種を駆逐してしまいます。[ 155 ]
キーストーン種とは、生態系の食物網の中で、その数に不釣り合いなほど大きな影響を及ぼす種のことである。[ 156 ]キーストーン種が除去されると、たとえバイオマスや生産性の尺度で生態系の小さな部分を占めていたとしても、生態系は劇的な変化を経験する可能性がある。[ 157 ]ラッコはウニがケルプの森に与えるダメージを抑えている。北米西海岸のラッコが毛皮目的で商業的に狩猟されたことで、その数があまりにも少なくなり、ウニの個体数を制御できなくなった。ウニは次に、ケルプの根元を大量に食べ尽くしたため、ケルプの森は、それに依存するすべての種とともに、ほぼ消滅した。ラッコを再導入することで、ケルプの生態系を回復することができた。[ 158 ] [ 159 ]
位相的位置
栄養段階間の相互作用のネットワークは、海洋生態系の機能について多くの情報を提供します。摂食習慣に加えて、様々な生物の3つの特性(移動性、サイズ、生息地)が、この栄養段階の見解を補完します。[ 160 ]

生態系の適切な機能を維持するためには、1994年にロートンが投げかけたシンプルな問い「生態系において種は何をするのか?」をより深く理解する必要がある。[ 161 ]生態学的役割と食物網の位置は独立していないため、[ 162 ]どのような種がネットワーク上の様々な位置を占めているかという問いが必要である。[ 160 ]キーストーン種を特定しようとする最初の試み以来、[ 163 ] [ 164 ]食物網におけるそれらの位置づけには関心が寄せられてきた。[ 165 ] [ 166 ]最初は頂点捕食者であったと示唆され、その後、植物、草食動物、寄生動物としても示唆された。[ 167 ] [ 168 ]群集生態学と保全生物学の両方にとって、それらが複雑な栄養段階ネットワークのどこに位置しているかを知ることは有用であろう。[ 160 ]
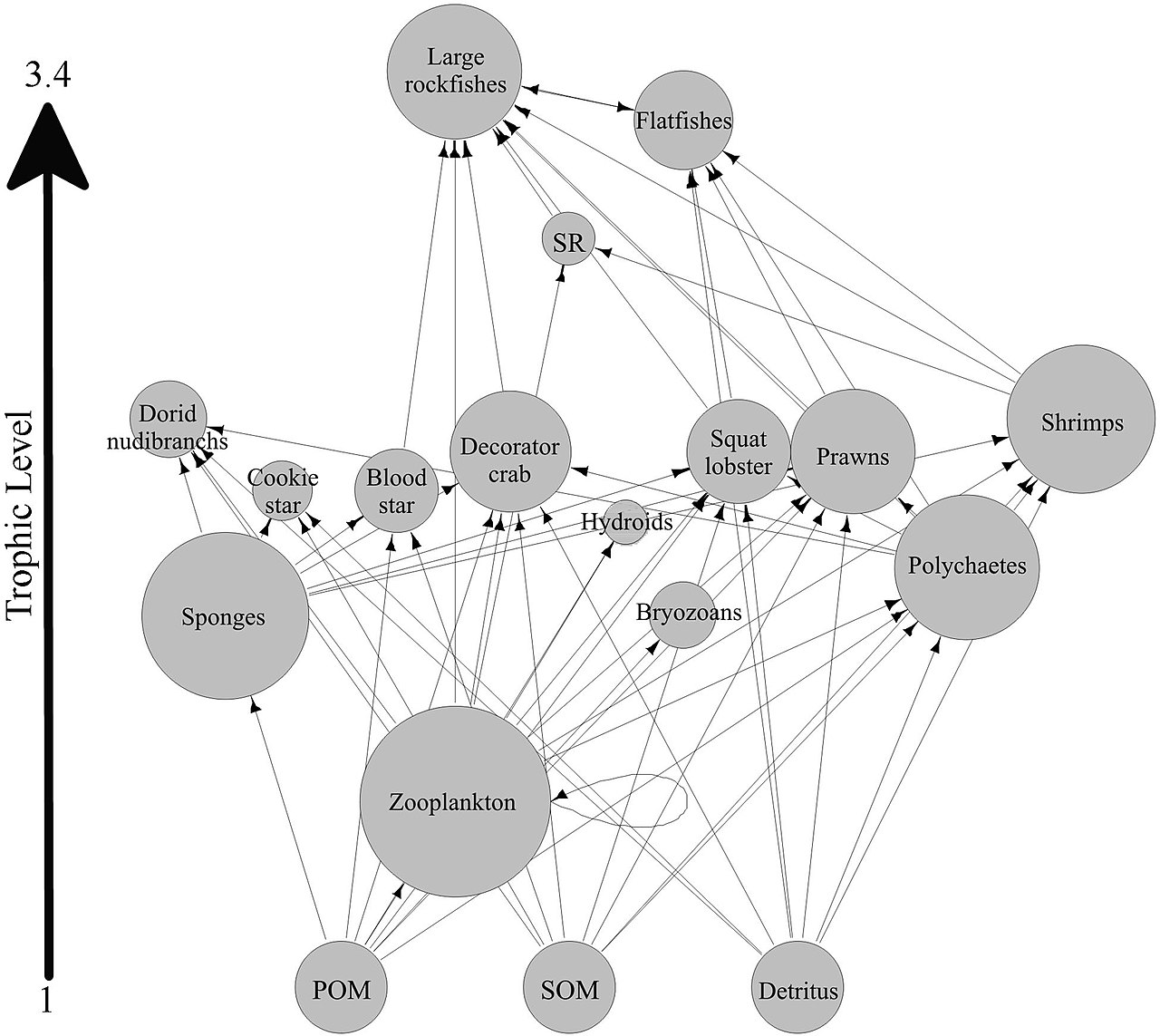
この種のネットワーク分析の一例は、海洋食物網のデータに基づいて図に示されている。[ 169 ]図は、食物網ノードの位相的位置と、関与する生物の移動度との関係を示している。食物網ノードは移動度に応じて形状コード化され、(A) ボトムアップグループ(固着性および漂流性)と (B) 食物網の頂点に位置するグループを強調する指標を用いて色分けされている。[ 160 ]
生物の相対的な重要性は時間と空間によって変化するため、大規模なデータベースを調べることで、問題に対する一般的な洞察が得られる可能性があります。異なる種類の生物が異なるタイプのネットワーク上の位置を占めている場合、食物網モデルにおいてこれを調整することで、より信頼性の高い予測が可能になります。中心性指標同士の比較(次数中心性と近接中心性の類似性、[ 170 ]、キーストーン指標とキーストーンネス指標、[ 171 ]、中心性指標と栄養段階(中程度の栄養段階にある高中心性の種の大部分)[ 172 ])は、食物網における生物の決定的に重要な位置をよりよく理解するために行われました。栄養段階に形質データを追加することでこの関心を広げると、結果の生物学的解釈に役立ちます。中心性指標間の関係は、生息地ネットワークなど、他のネットワークタイプでも研究されています。[ 173 ] [ 174 ]大規模なデータベースと新しい統計分析により、このような疑問を再調査し、知識を更新することができます。[ 160 ]
不可解なやり取り

海洋プランクトン食物網全体では、隠れた相互作用、つまり「目に見えない」相互作用が見られますが、現在、既存の手法ではほとんど見過ごされており、これらの相互作用に関する大規模なデータ収集は限られています。しかしながら、現在の証拠は、これらの相互作用の一部が食物網の動態やモデル結果に顕著な影響を及ぼす可能性を示唆しています。隠れた相互作用をモデルに組み込むことは、栄養素やエネルギーの輸送に関わる相互作用において特に重要です。[ 175 ]
この図は、5つの潜在的相互作用(混合栄養性、個体発生および種の違い、微生物の相互摂食、栄養要求性、細胞内炭素分配)によって影響を受ける物質フラックス、個体群、および分子プールを示しています。これらの相互作用は、食物網の領域が重複するため、相乗効果をもたらす可能性があります。例えば、植物プランクトンの細胞内炭素分配は、微生物の相互摂食に利用され、栄養要求性の場合は交換される下流の有機物プールと、個体発生および種の違いに基づく餌の選択の両方に影響を与える可能性があります。[ 175 ]
「動物プランクトンは植物プランクトンを消費する」「植物プランクトンは無機栄養素を吸収する」「総一次生産量は食物網に利用可能な炭素量を決定する」といった単純化は、科学者が水生環境における一般的な相互作用を説明し、モデル化するのに役立ってきました。従来の手法は、こうした一般化を定量化・定性化することに重点を置いていましたが、近年のゲノミクス、センサーの検出限界、実験手法、その他の技術の急速な進歩により、プランクトン群集内の相互作用を一般化することは単純すぎる可能性があることが示されました。こうした技術の向上により、バルクサンプリングの取り組みや実験手法によって不利な影響を受けているために不可解に見えていた多くの相互作用が明らかになりました。[ 175 ]
複雑さと安定性

食物網は、捕食者と被食者の複雑な相互作用ネットワークを構築するための枠組みを提供します。食物網モデルは、食物連鎖のネットワークです。各食物連鎖は、藻類や植物など、自ら食物を生産できる一次生産者または独立栄養生物から始まります。食物連鎖の次の生物は、一次生産者を餌とする生物であり、このようにして連鎖は捕食者の連鎖として続きます。各連鎖内の生物は、一次生産者から何リンク離れているかに基づいて、栄養段階に分類されます。食物連鎖の長さ、つまり栄養段階は、エネルギーまたは栄養素が植物から頂点捕食者へと移動する際に遭遇する種の数の尺度です。[ 178 ]食物エネルギーは、ある生物から次の生物へ、さらに次の生物へと流れ、各段階でいくらかのエネルギーが失われます。特定の栄養段階には、同じ捕食者と被食者を持つ1つの種または種群が存在する場合があります。[ 179 ]
1927年、チャールズ・エルトンは食物網の利用に関する影響力のある総合論文を発表し、これによって食物網は生態学における中心的な概念となった。[ 180 ] 1966年、ロバート・ペインが潮間帯の海岸についての実験的かつ記述的な研究を行い、食物網の複雑性が種の多様性と生態学的安定性の維持に重要であると示唆したことを受けて、食物網への関心が高まった。[ 181 ]ロバート・メイやスチュアート・ピムなど多くの理論生態学者が、この発見や他の発見に刺激を受けて食物網の数学的特性を調べた。彼らの分析によると、複雑な食物網は単純な食物網よりも安定性が低いはずである。[ 182 ] : 75–77 [ 183 ] : 64 自然界で観察される食物網の複雑さと食物網モデルの数学的脆弱性との間の明らかなパラドックスは、現在、集中的な研究と議論の対象となっている。このパラドックスは、食物網の持続性と食物網の平衡安定性の概念的な違いに部分的に起因している可能性がある。 [ 182 ] [ 183 ]
食物網内の栄養段階が抑制されると、食物網内で栄養カスケードが発生する可能性があり ます。
たとえば、捕食者が捕食において獲物の個体数を減らすか、またはその行動を変えるほど効果的である場合に、トップダウンのカスケードが発生し、それによって次の栄養段階が捕食から解放されます。トップダウンのカスケードは、頂点消費者/捕食者が一次消費者の個体数を管理する栄養カスケードです。その結果、一次生産者の個体数が繁栄します。頂点捕食者が除去されると、食物網のダイナミクスが変化する可能性があります。この場合、一次消費者は過剰に増殖し、一次生産者を搾取します。最終的には、消費者の個体数を維持するのに十分な一次生産者がいなくなります。トップダウンの食物網の安定性は、より高い栄養段階での競争と捕食に依存しています。外来種も、頂点捕食者を除去するか頂点捕食者になることで、このカスケードを変化させる可能性があります。この相互作用は必ずしも否定的とは限りません。研究によると、特定の外来種がカスケードをシフトさせ始めており、その結果、生態系の劣化が修復されています。[ 184 ] [ 185 ]複雑な外洋生態系におけるカスケードの一例は、1980年代から1990年代にかけて北大西洋で発生した。持続的な乱獲により大西洋タラ( Gadus morhua )やその他の底生魚が除去された結果、これらの底生魚の餌となる種、特にキタズワイガニ(Chionoecetes opilio)やキタエビ(Pandalus borealis )などの小型餌魚や無脊椎動物が増加した。これらの餌となる種の増加は、間接的な影響として、小型魚や無脊椎動物の餌となる動物プランクトンの群集を変化させた。 [ 186 ]トップダウンカスケードは、人間が狩猟や漁業を通じて多くの場所で行ってきたように、食物網から頂点捕食者を排除することの連鎖反応を理解する上で重要となりうる。
ボトムアップカスケードでは、一次生産者の個体群が常に高次栄養段階におけるエネルギーの増減を制御します。一次生産者は、光合成を必要とする植物、植物プランクトン、動物プランクトンです。光は重要ですが、一次生産者の個体群はシステム内の栄養素の量によって変化します。この食物網は、資源の利用可能性と限界に依存しています。すべての個体群は、最初に大量の栄養素が存在する場合、成長を経験します。[ 187 ] [ 188 ]
地上の比較

.jpg/1280px-Prochlorococcus_marinus_(cropped).jpg)
.jpg/1280px-Gnarly_Bristlecone_Pine_(crop).jpg)
海洋環境では、バイオマスピラミッドに逆転現象が生じることがあります。特に、消費者(コペポーダ類、オキアミ、エビ、餌となる魚類)のバイオマスは、一般的に一次生産者のバイオマスよりも大きいです。この逆転現象により、海洋動物のバイオマスの大部分は動物プランクトンによって構成されています。一次消費者である動物プランクトンは、一次生産者(主に植物プランクトン)と海洋食物網の残りの部分(二次消費者)をつなぐ重要な役割を担っています。[ 190 ]海洋の一次生産者のほとんどは、急速に成長し繁殖するというr戦略家の性質を持つ微小な植物プランクトンであるため、小さな塊でも一次生産速度が速いのです。
対照的に、成熟した森林などの多くの陸上一次生産者は、ゆっくりと成長し、再生産するK 戦略特性を持っているため、同じ一次生産率を達成するにははるかに大きな質量が必要です。生産率をそれを達成する平均バイオマス量で割ったものが、生物の生産量 / バイオマス (P/B) 比として知られています。[ 191 ]生産は、単位時間あたり面積あたりの質量またはエネルギーの移動量で測定されます。対照的に、バイオマスの測定は単位面積または体積あたりの質量の単位で行われます。P/B 比は、時間の逆単位 (例: 1/月) を使用します。この比率により、特定の栄養段階でのバイオマスの量と比較したエネルギーの流れの量を推定できるため、栄養段階間の区別が可能になります。P/B 比は、栄養段階と生物のサイズが増加するにつれて最も一般的には低下し、小さくて短命な生物は、大きくて長持ちする生物よりも高い P/B 比を含みます。
例:ブリスルコーンパインは数千年も生きることができますが、生産量/バイオマス比は非常に低いです。シアノバクテリアのプロクロロコッカスは24時間ほどしか生きられませんが、生産量/バイオマス比は非常に高いです。
海洋では、一次生産の大部分は藻類によって行われています。これは、一次生産の大部分が維管束植物によって行われている陸上とは対照的です。
| エコシステム | 純一次生産性(Gt /年) | 総植物バイオマス(Gt) | 回転時間(年) |
|---|---|---|---|
| 海洋 | 45~55歳 | 1~2 | 0.02~0.06 |
| 地上 | 55~70歳 | 600~1000 | 9~20 |

プランクトン性藻類や水生植物などの水生生産者には、陸上生態系の木本に見られるような二次成長の大きな蓄積がない。しかし、それらはより大きな草食動物のバイオマスを支えるのに十分な速さで繁殖することができる。これがピラミッドを逆転させる。一次消費者は寿命が長く成長速度が遅いため、消費する生産者よりも多くのバイオマスを蓄積する。植物プランクトンはわずか数日しか生きられないのに対し、植物プランクトンを食べる動物プランクトンは数週間生き、動物プランクトンを食べる魚は数年間連続して生きる。[ 194 ]水生捕食者は小型の消費者よりも死亡率が低い傾向があり、これが逆転したピラミッドのパターンに寄与している。個体群構造、移動速度、および獲物の環境的避難場所も、バイオマスが逆転したピラミッドの原因として考えられる。しかし、食物エネルギー源をすべて含めると、エネルギーピラミッドは常に直立したピラミッド型になる。これは熱力学の第二法則によるものである。」[ 195 ] [ 196 ]
生産された有機物のほとんどは、最終的には消費され、無機炭素に呼吸されます。堆積物に埋もれて保存される有機物の割合は、年間わずか2~4億トンで、総生産量のごく一部にすぎません。[ 54 ]世界の植物プランクトンの生産量は年間約500億トンで、植物プランクトンバイオマスは約10億トンであり、ターンオーバー時間は1週間です。海洋大型水草は、同様の世界のバイオマスを持っていますが、生産量は年間わずか10億トンであり、ターンオーバー時間は1年です。[ 197 ]これらの高いターンオーバー率(世界の陸上植生のターンオーバーが10~20年であるのと比較して)[ 192 ]は、有機物の安定した生産だけでなく、効率的な消費も意味します。有機物損失経路は複数存在する(自栄生物と従属栄養生物による呼吸、放牧、ウイルス溶解、デトリタス経路)が、いずれも最終的には呼吸と無機炭素の放出につながる。[ 54 ]
.jpeg)
人為的影響

- 乱獲
- 酸性化
翼足類とヒトデ類は共に北極の食物網の基盤を形成しており、どちらも酸性化によって深刻な被害を受けています。翼足類の殻は酸性化の進行とともに溶解し、ヒトデ類は付属肢の再生時に筋肉量を失います。[ 199 ]さらに、ヒトデ類の卵は、北極の酸性化に伴う予想される条件にさらされると数日以内に死滅します。[ 200 ]酸性化は北極の食物網を基盤から破壊する恐れがあります。北極の海水は急速に変化しており、アラゴナイトの飽和度が低下しつつあります。[ 201 ]北極の食物網は単純であると考えられており、小型生物から大型捕食者までの食物連鎖の段階は少ないことを意味します。例えば、翼足類は「大型プランクトン、魚類、海鳥、クジラなど、多くの高等捕食者にとって重要な獲物」です。[ 202 ]
.jpg/1280px-Fis01026_(27555118153).jpg)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
,_Belmont_-_geograph.org.uk_-_529175.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
- 気候変動
海洋生態系は、地球上の他のどの地域よりも気候変動の影響を受けやすい。これは、気温の上昇と海洋酸性化によるものである。海水温の上昇に伴い、魚類は既知の生息域から移動し、新たな生息域に定住すると予測されている。この変化の過程で、それぞれの種の個体数は大幅に減少する。現在、捕食者と被食者の間には、生存のために互いに依存している関係が数多く存在する。[ 203 ]種の生息域が変化すれば、捕食者と被食者の関係性や相互作用は大きな影響を受けるだろう。これらの変化が食物網のダイナミクスにどのような影響を与えるかを理解するための研究は現在も行われている。
科学者たちはモデリングを用いることで、特定の種が繁栄する地域や、その地域に生息する他の種との栄養段階的相互作用を分析することができます。最近のモデルでは、大型海洋生物の多くは、気候変動が示唆するよりも遅いペースで生息域を移動すると予測されています。これは捕食者と被食者の関係にさらに大きな影響を与えるでしょう。小型種や生物は、大型哺乳類よりも早く海洋の温暖化や移動の影響を受けやすいからです。[ 203 ]これらの捕食者は、小型種の移動に伴い、移動前に過去の生息域に長く留まることが分かっています。「新しい」種が大型哺乳類の生息域に侵入することで、生態系は変化し、捕食する獲物が増えます。[ 203 ]小型種の生息域は狭まる一方で、大型哺乳類は生息域を拡大するでしょう。こうした動態の変化は、海洋に生息するすべての種に大きな影響を与え、私たちの生態系全体にさらに多くの変化をもたらすでしょう。海の中で捕食者が獲物を見つけることができる場所の移動は、漁業にも影響を与えるでしょう。[ 204 ]現在、漁師は特定の魚種がどこに生息しているかを知っていますが、変化が起こると、漁師がどこで時間を過ごしているかを把握することが難しくなり、遠くまで移動する必要があるため、より多くのお金がかかります。[ 205 ]その結果、これらの魚の個体群の移動に伴って、特定の地域に設定されている現在の漁業規制に影響を及ぼす可能性があります。
.png)
プリンストン大学で実施された調査で、研究者たちは海洋生物が「気候速度」、つまり海洋生物の移動速度と方向と常に歩調を合わせていることを発見しました。1968年から2011年までのデータを調べたところ、動物の生息深度の変化の70%と緯度の変化の74%が、地域規模の海水温の変動と相関していることがわかりました。[ 208 ]これらの動きにより、生物は10年ごとに赤道から4.5マイルから40マイル(約72kmから84km)遠ざかっています。モデルの助けを借りれば、地域は生物がどこに行き着くかを予測できます。気候が生物に与える影響についての理解が深まるにつれて、モデルは変化に適応していく必要があります。
「私たちの研究結果は、将来の気候変動が、高次栄養段階へのエネルギーの流れの減少と、よりデトリタスベースのシステムへの移行を通じて海洋食物網を弱体化させる可能性があることを示しており、食物網の単純化と生産者と消費者のダイナミクスの変化につながり、どちらも底生生物群集の構造に重要な意味を持つ。」[ 209 ] [ 210 ]
「…気温上昇は、海洋食物網の底層にいる一次食物生産者(例えば藻類)から中間消費者(草食動物)、そして頂点にいる捕食者へと送られる重要なエネルギーの流れを減少させます。このようなエネルギー伝達の混乱は、頂点捕食者の食物供給量の減少につながる可能性があり、ひいては食物網内の多くの海洋生物種に悪影響を及ぼす可能性があります…気候変動は植物の生産性を高めましたが、これは主にシアノバクテリア(小型藍藻類)の増殖によるものです」とウラー氏は述べました。「しかし、この一次生産性の向上は食物網を支えるものではありません。なぜなら、これらのシアノバクテリアは大部分が食用にはならず、草食動物には食べられないからです。地球温暖化の影響下で生態系がどのように機能するかを理解することは、生態学研究における課題です。海洋温暖化に関する研究のほとんどは、1つまたは少数の種を対象とした単純化された短期的な実験に基づいています。」[ 210 ]


参照
参考文献
- ^ Dunne, Jennifer A.; Williams, Richard J.; Martinez, Neo D. (2002年10月). 「食物網の構造とネットワーク理論:連結性と規模の役割」 . Proceedings of the National Academy of Sciences . 99 (20 ) : 12917– 12922. Bibcode : 2002PNAS...9912917D . doi : 10.1073/pnas.192407699 . ISSN 0027-8424 . PMC 130560. PMID 12235364 .
- ^ Dunne, Jennifer A (2005年12月22日). 「食物網のネットワーク構造」 . Pascual, Mercedes; Dunne, Jennifer A (編). 『生態学的ネットワーク』 . Oxford University Press, New York, NY. pp. 27– 92. doi : 10.1093/oso/9780195188165.003.0002 . ISBN 978-0-19-518816-5。
- ^ Karlson, Agnes ML; Gorokhova, Elena; Gårdmark, Anna; Pekcan-Hekim, Zeynep; Casini, Michele; Albertsson, Jan; Sundelin, Brita; Karlsson, Olle; Bergström, Lena (2020年2月). 「バルト海における消費者の生理学的状態と食物網構造および被食者の食料価値との関連性」 . Ambio . 49 ( 2): 391– 406. Bibcode : 2020Ambio..49..391K . doi : 10.1007/s13280-019-01201-1 . ISSN 0044-7447 . PMC 6965491. PMID 31168701 .
- ^ Odum, WE ; Heald, EJ (1975)「河口域マングローブ群落のデトリタスを基盤とした食物網」LE Cronin編『河口域研究』第1巻、265~286ページ。Academic Press、ニューヨーク。
- ^ Pimm, SL; Lawton, JH (1978). 「複数の栄養段階における摂食について」. Nature . 275 (5680): 542– 544. Bibcode : 1978Natur.275..542P . doi : 10.1038/275542a0 . S2CID 4161183 .
- ^ Pauly, D.; Palomares, ML (2005). 「海洋食物網の破壊:それは我々が考えていたよりもはるかに広範囲に及んでいる」(PDF) . Bulletin of Marine Science . 76 (2): 197– 211. 2013年5月14日時点のオリジナル(PDF)からアーカイブ。 2020年3月20日閲覧。
- ^ Cortés, E. (1999). 「サメの標準化された食餌構成と栄養段階」 ICES J. Mar. Sci . 56 (5): 707– 717. Bibcode : 1999ICJMS..56..707C . doi : 10.1006/jmsc.1999.0489 .
- ^ Pauly, D.; Trites, A.; Capuli, E.; Christensen, V. (1998). 「海洋哺乳類の食事構成と栄養段階」 . ICES J. Mar. Sci . 55 (3): 467– 481. Bibcode : 1998ICJMS..55..467P . doi : 10.1006/jmsc.1997.0280 .
- ^研究者らが初めてヒトの栄養段階を計算Phys.org 2013年12月3日
- ^ボンホモー, シルヴァン; ドゥブロカ, ローラン; ル・パプ, オリヴィエ; バルデ, ジュリアン; カプラン, デイヴィッド・M.; シャソ, エマニュエル; ニーブラス, アンヌ=エリス (2013年12月17日). 「世界の食物網と人間の栄養段階の食べ尽くし」 .米国科学アカデミー紀要. 110 (51): 20617– 20620. Bibcode : 2013PNAS..11020617B . doi : 10.1073/pnas.1305827110 . ISSN 0027-8424 . PMC 3870703. PMID 24297882 .
- ^ a bクロロフィルNASA地球観測所。2019年11月30日にアクセス。
- ^郭、若宇;梁、燕島。シン、ユウ。ワン・ロング。モウ、シャンリー。曹操、春傑。謝、ルイゼ。チャン・チュアンルン。ティエン、ジウェイ。張永裕(2018年9月26日)。「最も深い生物圏であるマリアナ海溝におけるピコおよびナノ植物プランクトン群集への洞察」。微生物学のフロンティア。9 2289。Frontiers Media SA。土井:10.3389/fmicb.2018.02289。ISSN 1664-302X。PMC 6168665。PMID 30319587。
- ^ Heinrichs, Mara E.; Mori, Corinna; Dlugosch, Leon (2020). Jungblut, Simon; Liebich, Viola; Bode-Dalby, Maya (編).異なる視点から解明された水生生物とその化学環境の複雑な相互作用. シュプリンガー・インターナショナル・パブリッシング. pp. 279– 297. doi : 10.1007/978-3-030-20389-4_15 . ISBN 978-3-030-20388-7。
- ^ Field, CB (1998年7月10日). 「生物圏の一次生産:陸上と海洋の構成要素の統合」 . Science . 281 (5374): 237– 240. Bibcode : 1998Sci...281..237F . doi : 10.1126/science.281.5374.237 . PMID 9657713 .
- ^ Mouw, Colleen B.; Barnett, Audrey; McKinley, Galen A.; Gloege, Lucas; Pilcher, Darren (2016年10月20日). 「衛星データと統合した全球海洋微粒子有機炭素フラックス」 .地球システム科学データ. 8 (2): 531– 541. Bibcode : 2016ESSD....8..531M . doi : 10.5194/essd-8-531-2016 . ISSN 1866-3508 . S2CID 54551421 .
- ^キャンベル、マイク(2011年6月22日)「炭素隔離における海洋プランクトンの役割」 EarthTimes . 2014年8月22日閲覧。
- ^なぜ私たちは海を気にかけるべきなのでしょうか?NOAA:国立海洋局。2020年1月7日更新。2020年3月1日閲覧。
- ^ Kettler GC, Martiny AC, Huang K, Zucker J, Coleman ML, Rodrigue S, Chen F, Lapidus A, Ferriera S, Johnson J, Steglich C, Church GM, Richardson P, Chisholm SW (2007年12月). 「プロクロロコッカスの進化における遺伝子獲得と喪失のパターンと意味」 . PLOS Genetics . 3 (12) e231. doi : 10.1371/journal.pgen.0030231 . PMC 2151091. PMID 18159947 .
- ^ Nemiroff, R.; Bonnell, J. 編 (2006年9月27日). 「土星から見た地球」 . Astronomy Picture of the Day . NASA .
- ^ Partensky F, Hess WR, Vaulot D (1999年3月). 「プロクロロコッカス、世界的に重要な海洋光合成原核生物」 . Microbiology and Molecular Biology Reviews . 63 (1): 106–27 . doi : 10.1128/MMBR.63.1.106-127.1999 . PMC 98958. PMID 10066832 .
- ^ 「あなたが聞いたことのない最も重要な微生物」npr.org。
- ^ Mann, DG (1999). 「珪藻類における種概念」. Phycologia . 38 (6): 437– 495. Bibcode : 1999Phyco..38..437M . doi : 10.2216/i0031-8884-38-6-437.1 .
- ^ Lynn DH (2008年6月24日).繊毛原生動物:特性評価、分類、文献ガイド(第3版). Springer. p. 122. ISBN 978-1-4020-8239-9。
- ^ Copepodsの生物学Archived 2009-01-01 at the Wayback Machine at Carl von Ossietzky University of Oldenburg
- ^ a b c d Hays, Graeme C.; Doyle, Thomas K.; Houghton, Jonathan DR (2018年11月). 「クラゲの栄養段階における重要性のパラダイムシフト?」 . Trends in Ecology & Evolution . 33 (11): 874– 884. Bibcode : 2018TEcoE..33..874H . doi : 10.1016/j.tree.2018.09.001 . PMID 30245075 .
- ^ a bハミルトン、ギャリー(2016年3月24日). 「クラゲの秘密の生活」 . Nature . 531 (7595): 432– 434. Bibcode : 2016Natur.531..432H . doi : 10.1038/531432a . ISSN 0028-0836 . PMID 27008951 .
- ^カルドナ、ルイス;アルバレス・デ・ケベド、イレーネ。アサンプシオ州ボレル。アギラール、アレックス(2012年3月21日)。 Ropert-Coudert、Yan (編)。「地中海の頂点捕食者によるゼラチン状プランクトンの大量消費」。プロスワン。7 (3) e31329。Bibcode : 2012PLoSO...731329C。土井:10.1371/journal.pone.0031329。ISSN 1932-6203。PMC 3310041。PMID 22470416。
- ^海洋食物網の底辺に生息する小さな餌魚が新たな保護を受けるNational Public Radio、2016年4月7日。
- ^ Roman, J.; McCarthy, JJ (2010). 「クジラのポンプ:海洋哺乳類は沿岸流域における一次生産性を高める」 . PLOS ONE . 5 ( 10) e13255. Bibcode : 2010PLoSO...513255R . doi : 10.1371/journal.pone.0013255 . PMC 2952594. PMID 20949007. e13255.
- ^ブラウン、ジョシュア・E. (2010年10月12日). 「クジラの糞が海洋の健康状態を向上」 .サイエンス・デイリー. 2014年8月18日閲覧。
- ^ Raina, Jean-Baptiste (2018年4月24日). 「マイクロスケールの水生生物」 . mSystems . 3 (2) e00150-17. doi : 10.1128/mSystems.00150-17 . ISSN 2379-5077 . PMC 5881019. PMID 29629412 . 変更されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International Licenseに基づいて利用可能です。
- ^ Marco, D. 編 (2011).メタゲノミクス:現在のイノベーションと将来の動向. Caister Academic Press . ISBN 978-1-904455-87-5。
- ^ a b c Roslin, Tomas; Majaneva, Sanna (2016). 「食物網構築におけるDNAバーコードの利用 ― 陸生生態学者と水生生態学者が団結!」ゲノム59 ( 9 ): 603– 628. doi : 10.1139/gen-2015-0229 . PMID 27484156 . 。
- ^ Wilhelm, Steven W.; Suttle, Curtis A. (1999). 「海におけるウイルスと栄養循環」 . BioScience . 49 (10): 781– 788. doi : 10.2307/1313569 . ISSN 1525-3244 . JSTOR 1313569 .
- ^ Weinbauer, Markus G.; Hornák, Karel; Jezbera, Jan; Nedoma, Jiri; Dolan, John R.; Simek, Karel (2007年3月). 「ウイルス溶解と原生生物の放牧による細菌バイオマス、生産、多様性への相乗的および拮抗的影響」. Environmental Microbiology . 9 (3): 777– 788. Bibcode : 2007EnvMi...9..777W . doi : 10.1111/j.1462-2920.2006.01200.x . ISSN 1462-2912 . PMID 17298376 .
- ^ロビンソン、キャロル、ナガッパ・ラマイア。「微生物の従属栄養代謝速度は微生物の炭素ポンプを制約する。」アメリカ科学振興協会、2011年。
- ^ Van den Meersche、カレル;ミデルバーグ、ジャック J.カーライン・ステルト;ファン・ライスワイク、ピーター。ボシュカー、ヘンリカスTS;ヒープ、カルロ HR (2004 年 5 月)。「実験ブルーム中の炭素窒素カップリングと藻類と細菌の相互作用: 13 C トレーサー実験のモデル化」。陸水学と海洋学。49 (3): 862–878。ビブコード: 2004LimOc..49..862V。土井: 10.4319/lo.2004.49.3.0862。ISSN 0024-3590。
- ^ Käse, Laura; Geuer, Jana K. (2018). 「海洋気候変動に対する植物プランクトンの反応 ― 序論」. YOUMARES 8 – 境界を越えた海洋:互いに学び合う. pp. 55– 71. doi : 10.1007/978-3-319-93284-2_5 . ISBN 978-3-319-93283-5。修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International License に基づいて利用可能です。
- ^ハインリッヒス、マラ・E.、モリ、コリーナ、ドゥルゴッシュ、レオン (2020). 「水生生物とその化学環境の複雑な相互作用を様々な観点から解明」YOUMARES 9 - 海洋:私たちの研究、私たちの未来. pp. 279– 297. doi : 10.1007/978-3-030-20389-4_15 . ISBN 978-3-030-20388-7。。
- ^ Duponchel, Sarah; Fischer, Matthias G. (2019). 「Viva lavidaviruses! Five features of virophages that parasitize gigant DNA viruses」 . PLOS Pathogens . 15 (3): e1007592. doi : 10.1371/journal.ppat.1007592 . PMC 6428243. PMID 30897185 .
{{cite journal}}: CS1 maint: 記事番号をページ番号として表示 (リンク)修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International License に基づいて利用可能です。 - ^ a b c Suttle, Curtis A. (2007). 「海洋ウイルス — 地球生態系における主要な役割」. Nature Reviews Microbiology . 5 (10): 801– 812. doi : 10.1038/nrmicro1750 . PMID 17853907 . S2CID 4658457 .
- ^ a b c Middelboe, Mathias; Brussaard, Corina (2017). 「海洋ウイルス:海洋生態系の重要プレイヤー」 .ウイルス. 9 (10 ) : 302. Bibcode : 2017Virus...9..302M . doi : 10.3390/v9100302 . PMC 5691653. PMID 29057790 .
- ^バーグ、オーヴィンド;ボルシャイム、クヌート・イングヴェ。ブラットバック、グンナール。ヘルダル、ミカル (1989)。 「水生環境には大量のウイルスが存在する」。自然。340 (6233): 467–468 . Bibcode : 1989Natur.340..467B。土井:10.1038/340467a0。PMID 2755508。S2CID 4271861。
- ^ Roux, Simon; Hallam, Steven J.; Woyke, Tanja; Sullivan, Matthew B. (2015). 「公開されている微生物ゲノムから解明されたウイルスのダークマターとウイルス–宿主相互作用」 . eLife . 4 e08490 . doi : 10.7554/eLife.08490 . PMC 4533152. PMID 26200428 .
- ^ a b c d e Yau, Sheree; Seth-Pasricha, Mansha (2019). 「極地水生環境のウイルス」 .ウイルス. 11 ( 2): 189. Bibcode : 2019Virus..11..189Y . doi : 10.3390/v11020189 . PMC 6410135. PMID 30813316 . 変更されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International Licenseに基づいて利用可能です。
- ^ Fischer, MG; Allen, MJ; Wilson, WH; Suttle, CA (2010). 「注目すべき遺伝子群を持つ巨大ウイルスが海洋動物プランクトンに感染する」 ( PDF) . Proceedings of the National Academy of Sciences . 107 (45): 19508– 19513. Bibcode : 2010PNAS..10719508F . doi : 10.1073/pnas.1007615107 . PMC 2984142. PMID 20974979 .
- ^ Matthias G. Fischer; Michael J. Allen; William H. Wilson; Curtis A. Suttle (2010). 「注目すべき遺伝子群を持つ巨大ウイルスが海洋動物プランクトンに感染する」 ( PDF) . Proceedings of the National Academy of Sciences . 107 (45): 19508– 19513. Bibcode : 2010PNAS..10719508F . doi : 10.1073/pnas.1007615107 . PMC 2984142. PMID 20974979 .
- ^ Massana, Ramon; Javier Del Campo; Christian Dinter; Ruben Sommaruga (2007). 「ウイルス感染による海洋従属栄養性鞭毛藻Cafeteria roenbergensisの個体群減少」. Environmental Microbiology . 9 (11): 2660– 2669. Bibcode : 2007EnvMi...9.2660M . doi : 10.1111/ j.1462-2920.2007.01378.x . PMID 17922751. S2CID 30191542 .
- ^ Fischer MG, Suttle CA (2011年4月) . 「大型DNAトランスポゾンの起源となるヴィロファージ」. Science . 332 (6026): 231–4 . Bibcode : 2011Sci...332..231F . doi : 10.1126/science.11 99412. PMID 21385722. S2CID 206530677 .
- ^ Fischer MG, Hackl (2016年12月). 「宿主ゲノム統合と巨大ウイルスによるビロファージmavirusの再活性化」. Nature . 540 ( 7632): 288–91 . Bibcode : 2016Natur.540..288F . doi : 10.1038/nature20593 . PMID 27929021. S2CID 4458402 .
- ^加賀美麻衣子; 三木健; 滝本岳 (2014). 「Mycoloop:水生食物網におけるツボカビ」 . Frontiers in Microbiology . 5 : 166. doi : 10.3389/fmicb.2014.00166 . PMC 4001071. PMID 24795703 . 修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 3.0 International License に基づいて利用可能です。
- ^修正します、アンソニー。ブルゴー、ガエタン。カンリフ、マイケル。バージニア州エッジコム、P.エッティンガー、カサンドラ L.グティエレス、MH、ハイトマン、ジョセフ。ほら、エリック・FY。イアニリ、ジュゼッペ。ジョーンズ、アダム C.かがみ、まいこ。ピカード、キャスリン T.クワント、C. アリーシャ。ラグクマール、セシャギリ。リケルメ、メルティクセル;スタジッチ、ジェイソン。バルガス・ムニス、ホセ。ウォーカー、アリソン K.ヤーデン、オーデッド。グラドフェルター、エイミー S. (2019)。「海洋環境の菌類:未解決の疑問と未解決の問題」。mBio。10 (2) e01189-18。Bibcode : 2019mBio...1089.18A。土井:10.1128/mBio.01189-18。PMC 6401481。PMID 30837337。 修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International License に基づいて利用可能です。
- ^ Gutiérrez, Marcelo H.; Jara, Ana M.; Pantoja, Silvio (2016年5月). 「チリ中部沖のフンボルト海流系の湧昇生態系における真菌寄生虫による海洋珪藻類への感染」 . Environmental Microbiology . 18 (5): 1646– 1653. Bibcode : 2016EnvMi..18.1646G . doi : 10.1111/1462-2920.13257 . hdl : 10533/148260 . ISSN 1462-2912 . PMID 26914416 .
- ^ a b c d eミドルバーグ、ジャック・J. (2019). 「有機炭素から無機炭素への回帰」.海洋炭素生物地球化学. SpringerBriefs in Earth System Sciences. pp. 37– 56. doi : 10.1007/978-3-030-10822-9_3 . ISBN 978-3-030-10821-2. S2CID 104330175 .変更されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International Licenseに基づいて利用可能です。
- ^ Legendre L, Rassoulzadegan F (1995)「海洋水中のプランクトンと栄養塩の動態」オフィーリア、 41 :153–172。
- ^ Pace ML, Cole JJ, Carpenter SR, Kitchell JF (1999)「多様な生態系における栄養段階のカスケードの解明」 Trends Ecol Evol , 14 : 483–488.
- ^ Azam F, Fenchel T, Field JG, Gray JS, Meyer-Reil LA, Thingstad F (1983)「海における水柱微生物の生態学的役割」 Mar Ecol-Prog Ser , 10 : 257–263.
- ^ Anderson TRとLeB Williams PJ (1998)「イギリス海峡E1観測点における溶存有機炭素の季節サイクルのモデル化」 Estuar Coast Shelf Sci、 46 :93–109。
- ^ Van den Meersche K、Middelburg JJ、Soetaert K、van Rijswijk P、Boschker HTS、Heip CHR (2004) 「実験ブルーム中の炭素-窒素カップリングと藻類-細菌相互作用: 13C トレーサー実験のモデル化」。リムノル・オセアノグル、 49 : 862–878。
- ^ Suttle CA (2005)「海のウイルス」 Nature、 437 :356–361。
- ^ Siegel, David A.; Buesseler, Ken O.; Behrenfeld, Michael J.; Benitez-Nelson, Claudia R.; Boss, Emmanuel; Brzezinski, Mark A.; Burd, Adrian; Carlson, Craig A.; d'Asaro, Eric A.; Doney, Scott C.; Perry, Mary J.; Stanley, Rachel HR; Steinberg, Deborah K. (2016). 「世界の海洋純一次生産量の輸出と運命の予測:EXPORTS科学計画」 . Frontiers in Marine Science . 3 : 22. Bibcode : 2016FrMaS...3...22S . doi : 10.3389/fmars.2016.00022 .変更されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International Licenseに基づいて利用可能です。
- ^ Wang, Fuqiang; Wu, Ying; Chen, Zuozhi; Zhang, Guosen; Zhang, Jun; Zheng, Shan; Kattner, Gerhard (2019). 「安定同位体と脂肪酸で描いた南シナ海における中深海魚類の栄養段階間相互作用」 . Frontiers in Marine Science . 5 522. Bibcode : 2019FrMaS...5..522W . doi : 10.3389/fmars.2018.00522 .。
- ^ a b Tollefson, Jeff (2020). 「薄明帯へ:科学者たちが海洋の神秘の真ん中へ潜る」Nature . doi : 10.1038/d41586-020-00520-8 . PMID 33633390 . 。
- ^ a b Choy, C. Anela; Haddock, Steven HD; Robison, Bruce H. (2017). 「現場給餌観察による深海性食物網構造の解明」 . Proceedings of the Royal Society B: Biological Sciences . 284 (1868). Bibcode : 2017PBioS.28472116C . doi : 10.1098 / rspb.2017.2116 . PMC 5740285. PMID 29212727 . 修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International License に基づいて利用可能です。
- ^ Kelly, Thomas B.; Davison, Peter C.; Goericke, Ralf; Landry, Michael R.; Ohman, Mark D.; Stukel, Michael R. (2019). 「中深海食物網の維持における中型動物プランクトン日周垂直移動の重要性」 . Frontiers in Marine Science . 6 508. Bibcode : 2019FrMaS...6..508K . doi : 10.3389/fmars.2019.00508 .。
- ^深海の気候変動は今世紀半ばまでに7倍の速さになる可能性があるとガーディアン紙が報じている、2020年5月25日。
- ^ Brito-Morales, I., Schoeman, DS, Molinos, JG, Burrows, MT, Klein, CJ, Arafeh-Dalmau, N., Kaschner, K., Garilao, C., Kesner-Reyes, K. and Richardson, AJ (2020)「気候速度は深海生物多様性の将来の温暖化への曝露増加を明らかにする」 Nature Climate Change , pp.1-6. doi : 10.5281/zenodo.3596584 .
- ^イリゴエン、ザビエル; TA クレヴジャー;ロスタッド、A.アメリカ、マルティネス。ボイラ、G.アクーニャ、JL;ボード、A.エチェバリア、F.ゴンザレス・ゴルディージョ、ジリビア;ヘルナンデス・レオン、S.アグスティ、S.アクスネス、DL;ドゥアルテ、CM。カールトヴェット、S. (2014)。「外洋における大型中遠洋魚のバイオマスと栄養効率」ネイチャーコミュニケーションズ。5 3271. Bibcode : 2014NatCo...5.3271I。土井: 10.1038/ncomms4271。PMC 3926006。PMID 24509953。
- ^海洋の魚類バイオマスは推定値の10倍に上るEurekAlert、2014年2月7日。
- ^ Choy, CA; Wabnitz, CCC; Weijerman, M.; Woodworth-Jefcoats, PA; Polovina, JJ (2016). 「頂点への道を探る:海洋中栄養段階マイクロネクトン群の構成が北太平洋中央部における頂点捕食者のバイオマスを決定する仕組み」. Marine Ecology Progress Series . 549 : 9– 25. Bibcode : 2016MEPS..549....9C . doi : 10.3354/meps11680 .。
- ^ a b c d e倉田尚子; ケイト・ベラ; ブライアン・ハミルトン; マフムード・シブジ; アレクサンダー・ソロヴィエフ; シルビア・マット; オーレリアン・タルタル; ウィリアム・ペリー (2016). 「海洋表層付近におけるサーファクタント関連細菌」 . Scientific Reports . 6 19123. Bibcode : 2016NatSR...619123K . doi : 10.1038/ srep19123 . PMC 4709576. PMID 26753514 . 修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International License に基づいて利用可能です。
- ^ヴェラ・ハティッチ;チョソヴィッチ、ボエナ。マルチェンコ、エレナ。ビハリ、ネベンカ。フラノ・クルシニッチ(1981年)。 「海洋植物プランクトンによる界面活性剤の生産」。海洋化学。10 (6): 505–520。ビブコード: 1981MarCh..10..505Z。土井: 10.1016/0304-4203(81)90004-9。。
- ^ Levin, Lisa A.; Baco, Amy R.; Bowden, David A.; Colaco, Ana; Cordes, Erik E.; Cunha, Marina R.; Demopoulos, Amanda WJ; Gobin, Judith; Grupe, Benjamin M.; Le, Jennifer; Metaxas, Anna; Netburn, Amanda N.; Rouse, Greg W.; Thurber, Andrew R.; Tunnicliffe, Verena; Van Dover, Cindy Lee; Vanreusel, Ann; Watling, Les (2016). 「熱水噴出孔とメタン湧出帯:影響圏の再考」 . Frontiers in Marine Science . 3 : 72. Bibcode : 2016FrMaS...3...72L .土井:10.3389/fmars.2016.00072。
- ^ポルテイル、マリー;オル、カリン。デュボア、スタニスラス F.エスコバル=ブリオネス、エルバ。ジェリナス、イヴ。メノ、レナイク。ジョゼ・サラザン(2016)。「グアイマス盆地の熱水噴出孔と冷湧出部における食物網の複雑さ」。プロスワン。11 (9): e0162263。ビブコード: 2016PLoSO..1162263P。土井:10.1371/journal.pone.0162263。PMC 5040445。PMID 27683216。
{{cite journal}}: CS1 maint: 記事番号をページ番号として表示 (リンク)。 - ^ Bernardino, Angelo F.; Levin, Lisa A.; Thurber, Andrew R.; Smith, Craig R. (2012). 「噴出孔、湧水孔、有機物落下地点における堆積物マクロ動物相の比較構成、多様性、栄養生態」 . PLOS ONE . 7 (4): e33515. Bibcode : 2012PLoSO...733515B . doi : 10.1371/journal.pone.0033515 . PMID 22496753 .
{{cite journal}}: CS1 maint: 記事番号をページ番号として表示 (リンク)。 - ^ポーテイル、M.オル、K.エスコバー=ブリオネス、E.カプレイス、JC。メノット、L.ウェールズ、M.クルオード、P.サラディン首相。ゴッドフロイ、A.サラザン、J. (2015)。「グアイマス盆地の噴出孔と湧出部の大型動物群集の比較研究」。生物地球科学。12 (18): 5455–5479。Bibcode : 2015BGeo...12.5455P。土井:10.5194/bg-12-5455-2015。。
- ^ Harris, PT; Macmillan-Lawler, M.; Rupp, J.; Baker, EK (2014). 「海洋の地形学」.海洋地質学. 352 : 4– 24. Bibcode : 2014MGeol.352....4H . doi : 10.1016/j.margeo.2014.01.011 .
- ^ a b cオテロ、ショセ・ルイス;デ・ラ・ペーニャ・ラストラ、ソール。ペレスアルベルティ、アウグスト。フェレイラ、ティアゴ・オソリオ。ウエルタ=ディアス、ミゲル・アンヘル(2018)。 「窒素とリンの循環における重要な地球規模の推進力としての海鳥のコロニー」。ネイチャーコミュニケーションズ。9 246. Bibcode : 2018NatCo...9..246O。土井:10.1038/s41467-017-02446-8。PMID 29362437。 修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International Licenseに基づいて利用可能です。
- ^バレット, カイル; アンダーソン, ウェンディ B.; ウェイト, D. アレクサンダー; グリスマー, L. リー; ポリス†, ゲイリー A.; ローズ†, マイケル D. (2005). 「海洋補助金は島嶼部および沿岸部のトカゲ個体群の食生活と生息数を変化させる」.オイコス. 109 (1): 145– 153.書誌コード: 2005Oikos.109..145B . doi : 10.1111/j.0030-1299.2005.13728.x .
- ^ポリス, ゲイリー A.; ハード, スティーブン D. (1996). 「海洋と陸生食物網の連携:海洋からの外来生物の流入が小島嶼および沿岸陸域コミュニティにおける高い二次生産力を支える」.アメリカン・ナチュラリスト. 147 (3): 396– 423.書誌コード: 1996ANat..147..396P . doi : 10.1086/285858 . S2CID 84701185 .
- ^ジェンデ, スコット・M.; エドワーズ, リチャード・T.; ウィルソン, メアリー・F.; ウィプフリ, マーク・S. (2002). 「太平洋サケの水生および陸生生態系」 .バイオサイエンス. 52 (10): 917. doi : 10.1641/0006-3568(2002)052[0917:PSIAAT]2.0.CO;2 . ISSN 0006-3568 . JSTOR 10.1641/0006-3568(2002)052[0917:PSIAAT]2.0.CO;2 .
- ^ a bギャニオン、カリーヌ;ロトホイスラー、エヴァ。シルヤネン、アンネリ;イリ・レンコ、マリア。ヨルマライネン、ヴェイホ (2013)。「海鳥グアノはバルト海の沿岸食物網を肥沃にする」 .プロスワン。8 (4) e61284。Bibcode : 2013PLoSO...861284G。土井:10.1371/journal.pone.0061284。PMC 3623859。PMID 23593452。
- ^溝田千年; 登尾浩介; 森良明 (2012). 「中部日本における亜酸化窒素(N2O)排出の「ホットスポット」としてのカワウ(Phalacrocorax carbo)のコロニー」.大気環境. 57 : 29– 34. Bibcode : 2012AtmEn..57...29M . doi : 10.1016/j.atmosenv.2012.02.007 .
- ^ Bird, Michael I.; Tait, Elaine; Wurster, Christopher M.; Furness, Robert W. (2008). 「鳥類尿酸の安定炭素・窒素同位体分析」Rapid Communications in Mass Spectrometry . 22 (21): 3393– 3400. Bibcode : 2008RCMS...22.3393B . doi : 10.1002/rcm.3739 . PMID 18837063 .
- ^ a bコー、ステファン;アングロ、エレナ。ピサヌ、ブノワ。ルフィーノ、リセ。フォルキエ、ルーシー。ローヴェレック、オリヴィエ。シャピュイ、ジャン=ルイ。パスカル、ミシェル。ビダル、エリック。クールシャン、フランク (2012)。「島における同位体窒素の海鳥変調」。プロスワン。7 (6) e39125。Bibcode : 2012PLoSO...739125C。土井:10.1371/journal.pone.0039125。PMC 3377609。PMID 22723945。
- ^マルダー、クリスタ・PH、アンダーソン、ウェンディ・B、タウンズ、デイビッド・R、ベリンガム、ピーター・J(2011年9月8日)『海鳥諸島:生態、侵略、そして回復』Oup USA. ISBN 978-0-19-973569-3。
- ^ McFadden, Tyler Neal; Kauffman, J. Boone; Bhomia, Rupesh K. (2016). 「ホンジュラス、フォンセカ湾のマングローブにおける営巣水鳥の栄養レベルへの影響」 .湿地生態学と管理. 24 (2): 217– 229. Bibcode : 2016WetEM..24..217M . doi : 10.1007/s11273-016-9480-4 . S2CID 6021420 .
- ^エイドリアン・ズヴォリツキ;ズムドチンスカ・スカルベク、カタルジナ・マウゴルザタ。イリスコ、レッヒ。シュテンプニヴィッツ、レッヒ (2013)。「スピッツベルゲン島の板食性および魚食性の海鳥のコロニー付近におけるグアノの堆積と栄養豊富」。極地の生物学。36 (3): 363–372。ビブコード: 2013PoBio..36..363Z。土井:10.1007/s00300-012-1265-5。S2CID 12110520。
- ^ Doughty, Christopher E.; Roman, Joe; Faurby, Søren; Wolf, Adam; Haque, Alifa; Bakker, Elisabeth S.; Malhi, Yadvinder; Dunning, John B.; Svenning, Jens-Christian (2016). 「巨大生物の世界における全球栄養輸送」 . Proceedings of the National Academy of Sciences . 113 (4): 868– 873. Bibcode : 2016PNAS..113..868D . doi : 10.1073/ pnas.1502549112 . PMC 4743783. PMID 26504209 .
- ^ a b Honig, Susanna E.; Mahoney, Brenna (2016). 「ハワイ島オアフ島のサンゴ礁における海鳥のグアノ増加の証拠」. Marine Biology . 163 (2): 22. Bibcode : 2016MarBi.163...22H . doi : 10.1007/s00227-015-2808-4 . S2CID 87850538 .
- ^ Kolb, GS; Ekholm, J.; Hambäck, PA (2010). 「沿岸水域における海鳥の営巣コロニーによる藻類および水生無脊椎動物への影響」 .海洋生態学進歩シリーズ. 417 : 287–300 . Bibcode : 2010MEPS..417..287K . doi : 10.3354/meps08791 .
- ^ Wainright, SC; Haney, JC; Kerr, C.; Golovkin, AN; Flint, MV (1998). 「アラスカ州ベーリング海プリビロフ諸島セントポールにおける海鳥グアノ由来窒素の陸生植物および海生植物による利用」. Marine Biology . 131 (1): 63– 71. Bibcode : 1998MarBi.131...63W . doi : 10.1007/s002270050297 . S2CID 83734364 .
- ^ Staunton Smith, J.; Johnson, CR (1995). 「人口密集サンゴ礁における海鳥と人間からの栄養摂取」 . Marine Ecology Progress Series . 124 : 189–200 . Bibcode : 1995MEPS..124..189S . doi : 10.3354/meps124189 .
- ^ a b cロレイン、アン;ファニー・ウルブレク。ベンゾーニ、フランチェスカ。バルジョン、ルーシー。トレンブレイ・ボイヤー、ローラ。メンケス、クリストフ。ギリキン、デイビッド P.ペイリ、クロード。ジョルダン、エルベ。ブサリー、ジェルマン。フェルヘイデン、アヌーク。ビダル、エリック (2017)。「海鳥は太平洋の離島の造礁サンゴに窒素を供給します。 」科学的報告書。7 (1): 3721。ビブコード: 2017NatSR...7.3721L。土井:10.1038/s41598-017-03781-y。PMC 5473863。PMID 28623288。S2CID 6539261。 変更されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International Licenseに基づいて利用可能です。
- ^ Connell, Des W. (2018年5月4日). 『熱帯水系における汚染』 CRC Press. ISBN 978-1-351-09277-7。
- ^ハッチャー、ブルース・ゴードン (1990). 「サンゴ礁の一次生産性:パターンとプロセスの階層」. Trends in Ecology & Evolution . 5 (5): 149– 155. Bibcode : 1990TEcoE...5..149H . doi : 10.1016/0169-5347(90)90221-X . PMID 21232343 .
- ^ Falkowski, Paul G.; Dubinsky, Zvy; Muscatine, Leonard; McCloskey, Lawrence (1993). 「共生サンゴにおける個体群制御」. BioScience . 43 (9): 606– 611. doi : 10.2307/1312147 . JSTOR 1312147 .
- ^ a b Marubini, F.; Davies, PS (1996). 「硝酸塩はサンゴにおける褐虫藻の個体密度を増加させ、骨格形成を低下させる」.海洋生物学. 127 (2): 319– 328. Bibcode : 1996MarBi.127..319M . doi : 10.1007/BF00942117 . S2CID 85085823 .
- ^マスカティン、L.(1990)「サンゴ礁の炭素とエネルギーフラックスにおける共生藻類の役割」、エコシステムワールド、 25:75–87。
- ^ Ferrier, M. Drew (1991). 「4種のイシサンゴによる溶存遊離アミノ酸の純吸収量」Coral Reefs . 10 (4): 183– 187. Bibcode : 1991CorRe..10..183F . doi : 10.1007/BF00336772 . S2CID 25973061 .
- ^ Furla, P.; Allemand, D.; Shick, JM; Ferrier-Pagès, C.; Richier, S.; Plantivaux, A.; Merle, PL; Tambutté, S. (2005). 「共生花虫綱:藻類と動物の生理的キメラ」 .統合比較生物学. 45 (4): 595– 604. doi : 10.1093/icb/45.4.595 . PMID 21676806 .
- ^ミルズ, マシュー・M.; リップシュルツ, フレドリック; シーベンス, ケネス・P. (2004). 「4種のイシサンゴによる粒子状物質の摂取とそれに伴う窒素吸収」.サンゴ礁. 23 (3): 311– 323. Bibcode : 2004CorRe..23..311M . doi : 10.1007/s00338-004-0380-3 . S2CID 13212636 .
- ^ Mills, MM; Sebens, KP (2004). 「3種のサンゴによる底生堆積物からの窒素の摂取と同化」.海洋生物学. 145 (6): 1097–1106 . Bibcode : 2004MarBi.145 .1097M . doi : 10.1007/s00227-004-1398-3 . S2CID 84698653 .
- ^ Houlbrèque, F.; Tambutté, E.; Richard, C.; Ferrier-Pagès, C. (2004). 「イシサンゴ類にとっての微小食餌の重要性」 . Marine Ecology Progress Series . 282 : 151– 160. Bibcode : 2004MEPS..282..151H . doi : 10.3354/meps282151 .
- ^ Ferrier-Pagès, C.; Witting, J.; Tambutté, E.; Sebens, KP (2003). 「天然動物プランクトン摂食がイシサンゴStylophora pistillataの組織と骨格の成長に及ぼす影響」Coral Reefs . 22 (3): 229– 240. doi : 10.1007/s00338-003-0312-7 . S2CID 44869188 .
- ^マルビニ, フランチェスカ; タケ, ブレンダ (1999). 「重炭酸塩添加はサンゴの成長を促進する」 .陸水学・海洋学. 44 (3): 716– 720. Bibcode : 1999LimOc..44..716M . doi : 10.4319/lo.1999.44.3.0716 . S2CID 83654833 .
- ^フェリエ・パジェス、C.;ルクレール、N.ジョベール、J.ペレグリ、SP (2000)。「サンゴの滲出液によるピコプランクトンおよびナノプランクトンの成長の促進」。水生微生物生態学。21 : 203–209。Bibcode : 2000AqME...21..203F。土井:10.3354/ame021203。
- ^ Renegar, DA; Riegl, BM (2005). 「栄養塩濃度の上昇と二酸化炭素分圧の上昇が大西洋イシサンゴAcropora cervicornisの成長率に及ぼす影響」 .海洋生態学進歩シリーズ. 293 : 69– 76. Bibcode : 2005MEPS..293...69R . doi : 10.3354/meps293069 .
- ^ピーターセン、イェンス・ジェルルフ;ホルマー、マリアンヌ。テルマンセン、メッテ;ハスラー、ベリット (2019)。 「二枚貝による栄養の抽出」。海産二枚貝の商品とサービス。 pp. 179–208。土井:10.1007/978-3-319-96776-9_10。ISBN 978-3-319-96775-2。
- ^ Raicevich, S.; Granzotto, A.; Pastres, R.; Giovanardi, O.; Pranovi, F.; Libralato, S. (2003). 「ヴェネツィアのラグーンにおける機械式ハマグリ浚渫:栄養段階質量収支モデルを用いた生態系への影響評価」.海洋生物学. 143 (2): 393– 403. doi : 10.1007/s00227-003-1072-1 .
- ^米国地質調査所(USGS)「第14章:水鳥の食物と生息地の変化」図14.1.チェサピーク湾生態系に関する米国地質調査所の科学的知見の統合と環境管理への影響。USGSサーキュラー1316。この記事には、パブリックドメインであるこの情報源からのテキストが組み込まれています。
- ^ Perry, MC, Osenton, PC, Wells-Berlin, AM, Kidwell, DM (2005)、「大西洋岸の海ダックにおける歴史的食習慣との関連における食物選択」[抄録]、Perry, MC、第2回北米海ダック会議、2005年11月7日~11日、メリーランド州アナポリス、プログラムおよび抄録、USGSパタクセント野生生物研究センター、メリーランド州、123ページ(105ページ)。
- ^ Bowser, A. Kirsten; Diamond, Antony W.; Addison, Jason A. (2013). 「ツノメドリからプランクトンへ:メイン湾北部における海鳥の食物連鎖のDNAに基づく分析」 . PLOS ONE . 8 (12): e83152. Bibcode : 2013PLoSO...883152B . doi : 10.1371/journal.pone.0083152 . PMC 3865145. PMID 24358258 .
{{cite journal}}: CS1 maint: 記事番号をページ番号として表示 (リンク) - ^ Leray, Matthieu; Meyer, Christopher P.; Mills, Suzanne C. (2015). 「サンゴ礁に生息する捕食性魚類のメタバーコーディングによる食性分析は、サンゴ礁に生息する共生魚が、高度に分割された一般食性魚類の食性にわずかながら寄与していることを示している」 . PeerJ . 3 : e1047. doi : 10.7717/peerj.1047 . PMC 4485734. PMID 26137428 .
{{cite journal}}: CS1 maint: 記事番号をページ番号として表示 (リンク)。 - ^ Coll, Marta; Schmidt, Allison; Romanuk, Tamara; Lotze, Heike K. (2011). 「異なる空間スケールにおける海草群集の食物網構造と人間の影響」 . PLOS ONE . 6 (7): e22591. Bibcode : 2011PLoSO...622591C . doi : 10.1371/journal.pone.0022591 . PMID 21811637 .
{{cite journal}}: CS1 maint: 記事番号をページ番号として表示 (リンク)修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International License に基づいて利用可能です。 - ^ Stat, Michael; Huggett, Megan J.; Bernasconi, Rachele; Dibattista, Joseph D.; Berry, Tina E.; Newman, Stephen J.; Harvey, Euan S.; Bunce, Michael (2017). 「eDNAを用いた生態系バイオモニタリング:熱帯海洋環境における生命樹全体のメタバーコーディング」. Scientific Reports . 7 (1) 12240. Bibcode : 2017NatSR...712240S . doi : 10.1038/s41598-017-12501-5 . PMC 5612959. PMID 28947818 . 。
- ^ Archer, Stephanie K.; Kahn, Amanda S.; Thiess, Mary; Law, Lauren; Leys, Sally P.; Johannessen, Sophia C.; Layman, Craig A.; Burke, Lily; Dunham, Anya (2020年9月24日). 「基礎種の豊富さがガラススポンジ礁の食物網トポロジーに影響を与える」 . Frontiers in Marine Science . 7 549478. Frontiers Media SA. Bibcode : 2020FrMaS...749478A . doi : 10.3389/fmars.2020.549478 . ISSN 2296-7745 . 変更されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International Licenseに基づいて利用可能です。
- ^ロボ、エドゥアルド A.;ハインリヒ、カーラ・ギゼルダ。シュッハ、マリリア。ウェッツェル、カルロス・エドゥアルド。 Ector、Luc (2016)、Necchi JR、Orlando (編)、「河川における生物指標としての珪藻」、River Algae、Springer International Publishing、pp. 245–271、doi : 10.1007/978-3-319-31984-1_11、ISBN 978-3-319-31983-4
{{citation}}: CS1 maint: ISBNによる作業パラメータ(リンク) - ^ Stevenson, R. Jan; Pan, Yangdong; van Dam, Herman (2010), Smol, John P.; Stoermer, Eugene F. (eds.), "Assessing Environmental Conditions in rivers and streams with diatoms", The Diatoms (2 ed.), Cambridge University Press, pp. 57– 85, doi : 10.1017/cbo9780511763175.005 , ISBN 978-0-511-76317-5
{{citation}}: CS1 maint: ISBNによる作業パラメータ(リンク) - ^ McCarthy, JJ, Canziani, OF, Leary, NA, Dokken, DJ, White, KS (編) (2001)『気候変動2001:影響、適応、脆弱性:気候変動に関する政府間パネル第3次評価報告書に対する作業部会IIの貢献』 807ページ、ケンブリッジ大学出版局。ISBN 9780521015004
- ^ Murphy, EJ; Cavanagh, RD; Drinkwater, KF; Grant, SM; Heymans, JJ; Hofmann, EE; Hunt, GL; Johnston, NM (2016). 「極地外洋生態系の構造と機能の理解による変化の影響の予測」 . Proceedings of the Royal Society B: Biological Sciences . 283 (1844). Bibcode : 2016PBioS.28361646M . doi : 10.1098 / rspb.2016.1646 . PMC 5204148. PMID 27928038 .
- ^気候変動による生態系への影響:食物網の混乱EPA . 2020年2月11日にアクセス。この記事にはパブリックドメインのこの情報源からのテキストが組み込まれています。
- ^ 「北極海で『レジームシフト』が起こっていると科学者らは言う」 phys.org . 2020年8月16日閲覧。
- ^ Lewis, KM; Dijken, GL van; Arrigo, KR (2020年7月10日). 「植物プランクトン濃度の変化が北極海の一次生産の増加を促進」 . Science . 369 (6500): 198– 202. doi : 10.1126/science.aay8380 . ISSN 0036-8075 . PMID 32647002. S2CID 220433818. 2020年8月16日閲覧。
- ^ Stoecker, Diane K.; Lavrentyev, Peter J. (2018). 「極地海域の混合栄養性プランクトン:北極圏全域レビュー」 . Frontiers in Marine Science . 5 292. Bibcode : 2018FrMaS...5..292S . doi : 10.3389/fmars.2018.00292 .
- ^ Kilias, Estelle S.; Junges, Leandro; Šupraha, Luka; Leonard, Guy; Metfies, Katja; Richards, Thomas A. (2020). 「ツボカビの分布と珪藻類との共存は北極海の海氷融解と相関する」 . Communications Biology . 3 (1): 183. doi : 10.1038/s42003-020-0891-7 . PMC 7174370. PMID 32317738. S2CID 216033140 . 変更されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International Licenseに基づいて利用可能です。
- ^ Bender, Sara J.; Moran, Dawn M.; McIlvin, Matthew R.; Zheng, Hong; McCrow, John P.; Badger, Jonathan; Ditullio, Giacomo R.; Allen, Andrew E.; Saito, Mak A. (2018). 「 Phaeocystis antarcticaにおけるコロニー形成:分子メカニズムと鉄の生物地球化学との関連」 . Biogeosciences . 15 (16): 4923– 4942. Bibcode : 2018BGeo...15.4923B . doi : 10.5194/bg-15-4923-2018 . S2CID 92529531 .
- ^ Pinkernell, Stefan; Beszteri, Bánk (2014). 「気候変動が南極海の主要な珪酸塩沈降物の分布範囲に及ぼす潜在的影響」. Ecology and Evolution . 4 (16): 3147– 3161. Bibcode : 2014EcoEv...4.3147P . doi : 10.1002/ece3.1138 . PMC 4222203. PMID 25473469 .
- ^ Cavan, EL; Belcher, A.; Atkinson, A.; Hill, SL; Kawaguchi, S.; McCormack, S.; Meyer, B.; Nicol, S.; Ratnarajah, L.; Schmidt, K.; Steinberg, DK; Tarling, GA; Boyd, PW (2019). 「生物地球化学循環における南極オキアミの重要性」 . Nature Communications . 10 (1): 4742. Bibcode : 2019NatCo..10.4742C . doi : 10.1038/ s41467-019-12668-7 . PMC 6800442. PMID 31628346 . 修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International License に基づいて利用可能です。
- ^ a bコルドーネ、ジョージナ;マリーナ、トマス I.サリナス、ヴァネサ。ドイル、サンティアゴ R.サラビア、レオナルド A.モモ、フェルナンド R. (2018)。「南極海洋食物網における大型藻類損失の影響: 食物網研究への絶滅閾値の適用」。ピアJ。6 e5531。Bibcode : 2018PeerJ...6e5531C。土井:10.7717/peerj.5531。PMC 6139014。PMID 30225167。
- ^マリーナ、トマス I.サリナス、ヴァネサ。ジョージナ、コルドーネ。カンパーナ、ガブリエラ。モレイラ、ユージニア。デレギバス、ドロレス。トーレ、ルシアナ。サハデ、リカルド。タティアン、マルコス。バレラ・オロ、エステバン。デ・トロック、マーリーン。ドイル、サンティアゴ。クアルティーノ、マリア・リリアナ。サラビア、レオナルド A.モモ、フェルナンド R. (2018)。 「ポッターコーブ(南極大陸)の食物網:複雑さ、構造、機能」。河口、海岸、陸棚の科学。200 : 141–151。Bibcode : 2018ECSS..200..141M。doi : 10.1016/j.ecss.2017.10.015 . hdl : 11336/39918 .
- ^ Koh, Eileen Y.; Martin, Andrew R.; McMinn, Andrew; Ryan, Ken G. (2012). 「南極海氷における微生物光合成の最近の進歩と将来の展望」 .生物学. 1 (3): 542– 556. doi : 10.3390/biology1030542 . PMC 4009807. PMID 24832507 .
- ^
- Azam, F.; Fenchel, T.; Field, JG; Gray, JS; Meyer-Reil, LA; Thingstad, F. (1983). 「海における水柱微生物の生態学的役割」 .海洋生態学進歩シリーズ. 10 (3): 257– 263. Bibcode : 1983MEPS...10..257A . doi : 10.3354/meps010257 . JSTOR 24814647 .
- フェンチェル、トム (2008). 「微生物ループ ― 25年後」. Journal of Experimental Marine Biology and Ecology . 366 ( 1–2 ): 99–103 . Bibcode : 2008JEMBE.366...99F . doi : 10.1016/j.jembe.2008.07.013 .
- ^ a b c Boetius, Antje; Anesio, Alexandre M.; Deming, Jody W.; Mikucki, Jill A.; Rapp, Josephine Z. (2015). 「氷圏の微生物生態学:海氷と氷河生息地」 . Nature Reviews Microbiology . 13 (11): 677– 690. doi : 10.1038/nrmicro3522 . PMID 26344407. S2CID 20798336 .
- ^ランペロット、パブロ (2014). 「極地微生物学:最近の進歩と将来の展望」 .生物学. 3 (1): 81– 84. doi : 10.3390/biology3010081 . PMC 4009764 .
- ^ a b Laybourn-Parry, Johanna (2009). 「No Place Too Cold」. Science . 324 ( 5934): 1521– 1522. Bibcode : 2009Sci...324.1521L . doi : 10.1126/science.11 73645. PMID 19541982. S2CID 33598792 .
- ^ Cavicchioli, Ricardo (2015). 「南極水圏の微生物生態学」. Nature Reviews Microbiology . 13 (11): 691– 706. doi : 10.1038/nrmicro3549 . hdl : 1959.4/unsworks_49930 . PMID 26456925. S2CID 23089203 .
- ^ a b Anesio, Alexandre M.; Bellas, Christopher M. (2011). 「低温生息地はウイルスによって引き起こされる微生物進化のホットスポットか?」Trends in Microbiology . 19 (2): 52– 57. Bibcode : 2011TrMic..19...52A . doi : 10.1016/j.tim.2010.11.002 . PMID 21130655 .
- ^ a b Anesio, Alexandre M.; Laybourn-Parry, Johanna (2012). 「バイオームとしての氷河と氷床」. Trends in Ecology & Evolution . 27 (4): 219– 225. Bibcode : 2012TEcoE..27..219A . doi : 10.1016/j.tree.2011.09.012 . PMID 22000675 .
- ^ a b c d eギリオーネ、J.-F.;ガーランド、PE;ポミエ、T.ペドロス・アリオ、C.マース、EW;バッカー、K.バーティルソン、S. DL州キルヒマン。ラブジョイ、C.イェーガー、PL;マレー、オーストラリア (2012)。「海洋表層および深層の細菌群集の極から極までの生物地理学」。米国科学アカデミーの議事録。109 (43): 17633–17638。Bibcode : 2012PNAS..10917633G。土井:10.1073/pnas.1208160109。PMC 3491513。PMID 23045668。
- ^ Herron, Ellen M.; Dewey, John F.; Pitman, WC (1974). 「北極圏の進化に関するプレートテクトニクスモデル」. Geology . 2 (8): 377. Bibcode : 1974Geo.....2..377H . doi : 10.1130/0091-7613(1974)2<377:PTMFTE>2.0.CO;2 . ISSN 0091-7613 .
- ^ Gaina, Carmen; Medvedev, Sergei; Torsvik, Trond H.; Koulakov, Ivan; Werner, Stephanie C. (2014). 「4D北極:新たな地球物理地図、プレートテクトニクス、トモグラフィーモデルに基づく北極圏の構造と進化の概観」. Surveys in Geophysics . 35 (5): 1095– 1122. Bibcode : 2014SGeo...35.1095G . doi : 10.1007/s10712-013-9254-y . PMC 4456077. PMID 26069354 .
- ^金尾正樹; スヴォーロフ, ウラジミール・D.; 戸田茂; 坪井誠治 (2015). 「北極圏における地震活動、構造、テクトニクス」 .地球科学フロンティア. 6 (5): 665– 677. Bibcode : 2015GeoFr...6..665K . doi : 10.1016/j.gsf.2014.11.002 . S2CID 129578736 .
- ^ Adie, RJ (1962)「南極の地質学」『南極研究:マシュー・フォンテーン・モーリー記念シンポジウム』John Wiley & Sons.
- ^ Baldwin, AJ; Moss, JA; Pakulski, JD; Joux, F.; Jeffrey, WH (2005). 「北極圏から南極圏に至る太平洋横断帯における微生物多様性」 .水生微生物生態学. 41 : 91–102 . doi : 10.3354/ame041091 .
- ^ロペス=ブエノ、アルベルト;タメス、ハビエル。ベラスケス、デイビッド。モヤ、アンドレス。ケサダ、アントニオ。アルカミ、アントニオ (2009)。 「南極の湖からのウイルスコミュニティの多様性」。科学。326 (5954): 858–861 . Bibcode : 2009Sci...326..858L。土井:10.1126/science.1179287。PMID 19892985。S2CID 32607904。
- ^ Säwström, Christin; Lisle, John; Anesio, Alexandre M.; Priscu, John C.; Laybourn-Parry, Johanna (2008). 「極地内水域におけるバクテリオファージ」 .極限環境微生物. 12 (2): 167– 175. Bibcode : 2008Extmo..12..167S . doi : 10.1007/ s00792-007-0134-6 . PMID 18188502. S2CID 2927907 .
- ^ Lamy, Thomas; Koenigs, Craig; Holbrook, Sally J.; Miller, Robert J.; Stier, Adrian C.; Reed, Daniel C. (2020). 「基盤種はジャイアントケルプ林における多様性増加によって群集の安定性を促進する」. Ecology . 101 (5) e02987. Bibcode : 2020Ecol..101E2987L . doi : 10.1002/ecy.2987 . PMID 31960414 . 。
- ^ Dayton, PK 1972.南極マクマード湾におけるコミュニティの回復力とエンリッチメントの底生生物への潜在的影響の理解に向けて。pp. 81–96、保全問題に関するコロキウム議事録、アレンプレス、ローレンス、カンザス州。
- ^ジャイアントケルプは南カリフォルニアの海洋生態系に強固な基盤を与える、国立科学財団、2020年2月4日。
- ^アンジェリーニ, クリスティン; アルティエリ, アンドリュー H.; 他 (2011年10月). 「基礎種間の相互作用とそれが群集組織、生物多様性、そして保全に及ぼす影響」 .バイオサイエンス. 61 (10): 782– 789. Bibcode : 2011BiSci..61..782A . doi : 10.1525/bio.2011.61.10.8 .
- ^ Ellison, Aaron M.; Bank, Michael S.; et al. (2005年11月). 「基盤種の喪失:森林生態系の構造と動態への影響」 . Frontiers in Ecology and the Environment . 3 (9): 479– 486. doi : 10.1890/1540-9295(2005)003[0479:LOFSCF]2.0.CO;2 . hdl : 11603/29165 .
- ^ペイン, RT (1969). 「栄養段階の複雑性と群集の安定性に関する覚書」.アメリカンナチュラリスト. 103 (929): 91– 93. Bibcode : 1969ANat..103...91P . doi : 10.1086/282586 . JSTOR 2459472. S2CID 83780992 .
- ^ 「キーストーン種仮説」ワシントン大学。2011年1月10日時点のオリジナルよりアーカイブ。2011年2月3日閲覧。
- ^ペイン, RT (1966). 「食物網の複雑性と種の多様性」.アメリカンナチュラリスト. 100 (910): 65– 75. Bibcode : 1966ANat..100...65P . doi : 10.1086/282400 . JSTOR 2459379. S2CID 85265656 .
- ^ペイン, RT (1995). 「キーストーン種の概念の洗練に関する対話」.保全生物学. 9 (4): 962– 964. Bibcode : 1995ConBi...9..962P . doi : 10.1046/j.1523-1739.1995.09040962.x .
- ^ Davic, Robert D. (2003). 「キーストーン種と機能グループの関連付け:キーストーン種概念の新たな操作的定義」 . Conservation Ecology . 7 (1) resp11. Bibcode : 2003ConEc...7Pes11D . doi : 10.5751/ES-00502-0701r11 . 2003年8月26日時点のオリジナルよりアーカイブ。2011年2月3日閲覧。
- ^ Szpak, Paul; Orchard, Trevor J.; Salomon, Anne K.; Gröcke, Darren R. (2013). 「ハイダ・グワイ南部(カナダ、ブリティッシュコロンビア州)における地域的生態学的変動と海上毛皮貿易による沿岸生態系への影響:メバル(Sebastes spp.)骨コラーゲンの安定同位体分析による証拠」 . Archaeological and Anthropological Sciences . 5 (2): 159– 182. Bibcode : 2013ArAnS...5..159S . doi : 10.1007/s12520-013-0122-y . S2CID 84866250 .
- ^ Cohn, JP (1998). 「ラッコを理解する」 . BioScience . 48 (3): 151– 155. doi : 10.2307/1313259 . JSTOR 1313259 .
- ^ a b c d e f Endrédi, Anett; Patonai, Katalin; Podani, János; Libralato, Simone; Jordán, Ferenc (2021). 「海洋食物網における誰がどこにいるのか? 特性に基づくネットワークポジションの分析」 . Frontiers in Marine Science . 8 636042. Bibcode : 2021FrMaS...836042E . doi : 10.3389/fmars.2021.636042 . hdl : 10831/110681 .変更されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International Licenseに基づいて利用可能です。
- ^ロートン、ジョン・H. (1994). 「生態系における種の活動とは?」オイコス. 71 (3): 367– 374. Bibcode : 1994Oikos..71..367L . doi : 10.2307/3545824 . JSTOR 3545824 .
- ^ Luczkovich, Joseph J.; Borgatti, Stephen P.; Johnson, Jeffrey C.; Everett, Martin G. (2003). 「食物網における栄養役割の類似性の定義と測定:正規同値性を用いた」. Journal of Theoretical Biology . 220 (3): 303– 321. Bibcode : 2003JThBi.220..303L . doi : 10.1006/jtbi.2003.3147 . PMID 12468282 .
- ^ペイン, ロバート・T. (1966). 「食物網の複雑性と種の多様性」.アメリカン・ナチュラリスト. 100 (910): 65– 75. Bibcode : 1966ANat..100...65P . doi : 10.1086/282400 . S2CID 38675566 .
- ^ペイン, ロバート・T. (1969). 「ピサスター-テグラ相互作用:捕食者パッチ、捕食者の食物嗜好、そして潮間帯の群集構造」.生態学. 50 (6): 950–961 . Bibcode : 1969Ecol...50..950P . doi : 10.2307/1936888 . JSTOR 1936888 .
- ^ミルズ, L. スコット; ソウル, マイケル E.; ドーク, ダニエル F. (1993). 「生態学と保全におけるキーストーン種概念」 .バイオサイエンス. 43 (4): 219– 224. doi : 10.2307/1312122 . JSTOR 1312122 .
- ^メアリー・E・パワー、デイビッド・ティルマン、ジェームズ・A・エステス、ブルース・A・メンジ、ウィリアム・J・ボンド、L・スコット・ミルズ、グレッチェン・デイリー、フアン・カルロス・カスティーリャ、ジェーン・ルブチェンコ、ロバート・T・ペイン (1996). 「キーストーン探索における課題」 . BioScience . 46 (8): 609– 620. Bibcode : 1996BiSci..46..609P . doi : 10.2307/1312990 . JSTOR 1312990 .
- ^ Bond, WJ (1994). 「キーストーン種」.生物多様性と生態系機能. pp. 237– 253. doi : 10.1007/978-3-642-58001-7_11 . ISBN 978-3-540-58103-1。
- ^ Marcogliese, David J.; Cone, David K. (1997). 「食物網:寄生虫への嘆願」. Trends in Ecology & Evolution . 12 (8): 320– 325. Bibcode : 1997TEcoE..12..320M . doi : 10.1016/s0169-5347(97)01080-x . PMID 21238094 .
- ^ Pinnegar, John.K; Polunin, Nicholas VC (2004). 「動的シミュレーションモデルを用いた地中海の岩礁沿岸域における漁業の間接的影響の予測」 .生態学的モデリング. 172 ( 2–4 ): 249– 267. Bibcode : 2004EcMod.172..249P . doi : 10.1016/J.ECOLMODEL.2003.09.010 .
- ^ヨルダン、フェレンツ;ベネデク、ゾフィア。ポダーニ、ヤーノス (2007)。 「食物網における位置の重要性の定量化:中心性指数の比較」。生態学的モデリング。205 ( 1–2 ): 270–275。Bibcode : 2007EcMod.205..270J。土井:10.1016/j.ecolmodel.2007.02.032。
- ^エンドレディ、アネット;セナンスキー、ベラ。リブララート、シモーネ。フェレンツ、ヨルダン(2018)。 「栄養階層における食物網の動態」。生態学的モデリング。368 : 94–103。Bibcode : 2018EcMod.368...94E。土井:10.1016/j.ecolmodel.2017.11.015。
- ^ Scotti, M.; Jordán, F. (2010). 「食物網における中心性指標と栄養段階の関係」.群集生態学. 11 (1): 59– 67. Bibcode : 2010ComEc..11...59S . doi : 10.1556/comec.11.2010.1.9 .
- ^ Baranyi, Gabriella; Saura, Santiago; Podani, János; Jordán, Ferenc (2011). 「ネットワークの連結性に対する生息地パッチの寄与:位相指標の冗長性と一意性」.生態指標. 11 (5): 1301– 1310. Bibcode : 2011EcInd..11.1301B . doi : 10.1016/j.ecolind.2011.02.003 .
- ^ Pereira, Juliana; Saura, Santiago; Jordán, Ferenc (2017). 「景観グラフ分析における単一ノード中心性 vs. 複数ノード中心性:スペイン北東部における20種の鳥類にとっての重要な生息地パッチとその保護」 . Methods in Ecology and Evolution . 8 (11): 1458– 1467. Bibcode : 2017MEcEv...8.1458P . doi : 10.1111/2041-210X.12783 . S2CID 90319704 .
- ^ a b c d Millette, Nicole C.; Grosse, Julia; Johnson, Winifred M.; Jungbluth, Michelle J.; Suter, Elizabeth A. (2018). 「平凡な光景の中に隠されたもの:海洋プランクトンにおける潜在的相互作用の重要性」. Limnology and Oceanography Letters . 3 (4): 341– 356. Bibcode : 2018LimOL...3..341M . doi : 10.1002/lol2.10084 .修正されたテキストはこのソースからコピーされたもので、Creative Commons Attribution 4.0 International License に基づいて利用可能です。
- ^ a b cルイパート、トーマス;ヘイガン、ジェームス G.マッカーシー、モーガン L.ポティ、ミーナクシ (2020)。ユングブルート、サイモン。リービヒ、ヴィオラ。ボード・ダルビー、マヤ (編)。人新世の海洋生物多様性の状況。チャム:シュプリンガー・インターナショナル・パブリッシング。 pp. 57–82 .土井: 10.1007/978-3-030-20389-4_4。ISBN 978-3-030-20388-7。
- ^ Estes, JA; Tinker, MT; Williams, TM; Doak, DF (1998). 「シャチによるラッコの捕食:海洋生態系と沿岸生態系のつながり」. Science . 282 (5388): 473– 476. Bibcode : 1998Sci...282..473E . doi : 10.1126/science.282.5388.473 . PMID 9774274 . 。
- ^ Post, DM (1993). 「食物連鎖の長さの長所と短所」. Trends in Ecology and Evolution . 17 (6): 269– 277. doi : 10.1016/S0169-5347(02)02455-2 .
- ^ジェリー・ボブロウ博士、スティーブン・フィッシャー(2009年)『クリフスノーツ CSET:多科目対策(第2版)』ジョン・ワイリー・アンド・サンズ、283ページ。ISBN 978-0-470-45546-3。
- ^エルトン・CS(1927)『動物生態学』2001年再版、シカゴ大学出版局。
- ^ Paine RT (1966). 「食物網の複雑性と種の多様性」. The American Naturalist . 100 (910): 65– 75. Bibcode : 1966ANat..100...65P . doi : 10.1086/282400 . S2CID 85265656 .
- ^ a b RM 5月 (2001)モデル生態系における安定性と複雑性プリンストン大学出版局、1973年版の再版(新しい序文付き) 。ISBN 978-0-691-08861-7。
- ^ a b Pimm SL (2002) Food Webs University of Chicago Press、1982年版の再版(新しい序文付き) 。ISBN 978-0-226-66832-1。
- ^コッタ、J.;ウェルンバーグ、T.ジェーンズ、H.コッタ、I。ヌルクセ、K.ペルノハ、M.オラフ・コッタ、H. (2018)。「新しいカニの捕食者が海洋生態系の変化を引き起こす」 .科学的報告書。8 (1): 4956。Bibcode : 2018NatSR...8.4956K。土井:10.1038/s41598-018-23282-w。PMC 5897427。PMID 29651152。
- ^ Megrey, Bernard、Werner, Francisco. 「モデリングの観点から見たトップダウン型とボトムアップ型のエコシステム規制の役割の評価」(PDF)。
{{cite web}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ Frank, KT; Petrie, B.; Choi, JS; Leggett, WC (2005). 「かつてタラが優占していた生態系における栄養段階のカスケード」. Science . 308 (5728): 1621– 1623. Bibcode : 2005Sci...308.1621F . doi : 10.1126 / science.11 13075. ISSN 0036-8075 . PMID 15947186. S2CID 45088691 .
- ^松崎 真一郎 S.; 鈴木 健太; 角谷 拓; 中川 恵; 高村 典子 (2018). 「浅い高栄養湖における一次生産、動物プランクトン、魚類のボトムアップ連鎖」 .生態学. 99 ( 9): 2025– 2036. Bibcode : 2018Ecol...99.2025M . doi : 10.1002/ecy.2414 . PMID 29884987. S2CID 46996957 .
- ^ Lynam, Christopher Philip; Llope, Marcos; Möllmann, Christian; Helaouët, Pierre; Bayliss-Brown, Georgia Anne; Stenseth, Nils C. (2017年2月). 「北海における栄養段階と環境制御」 . Proceedings of the National Academy of Sciences . 114 (8): 1952– 1957. doi : 10.1073/pnas.1621037114 . PMC 5338359. PMID 28167770 .
- ^ 「Oldlist」 . ロッキー山脈年輪研究. 2013年1月8日閲覧。
- ^米国エネルギー省 (2008)「炭素循環と生物隔離」 81ページ、ワークショップ報告書 DOE/SC-108、米国エネルギー省科学局。
- ^ Wetzel, Robert (2001). Limnology: Lake and river ecosystems . San Diego: Academic Press. ISBN 978-0-12-744760-5。
{{cite book}}: CS1 maint: 発行者の所在地 (リンク) - ^ a b Field, CB; Behrenfeld, MJ; Randerson, JT; Falkowski, P. (1998). 「生物圏の一次生産:陸上と海洋の構成要素の統合」 . Science . 281 (5374): 237– 240. Bibcode : 1998Sci...281..237F . doi : 10.1126/science.281.5374.237 . PMID 9657713 .
- ^ Bar-On, Yinon M.; Phillips, Rob; Milo, Ron (2018). 「地球上のバイオマス分布」 . Proceedings of the National Academy of Sciences . 115 (25): 6506– 6511. Bibcode : 2018PNAS..115.6506B . doi : 10.1073/pnas.1711842115 . PMC 6016768. PMID 29784790 . 。
- ^スペルマン、フランク・R. (2008). 『水の科学:概念と応用』CRCプレス. p. 167. ISBN 978-1-4200-5544-3。
- ^ Odum, EP; Barrett, GW (2005). 『生態学の基礎』(第5版). Brooks/Cole, Cengage Learning. ISBN 978-0-534-42066-6. 2011年8月20日時点のオリジナルよりアーカイブ。
- ^ Wang, H.; Morrison, W.; Singh, A.; Weiss, H. (2009). 「生態系における逆バイオマスピラミッドと避難所のモデリング」(PDF) .生態学的モデリング. 220 (11): 1376– 1382. Bibcode : 2009EcMod.220.1376W . doi : 10.1016/j.ecolmodel.2009.03.005 . 2011年10月7日時点のオリジナル(PDF)からアーカイブ。 2020年3月18日閲覧。
- ^ Smith, SV (1981). 「地球規模の炭素吸収源としての海洋大型水草類」. Science . 211 (4484): 838– 840. Bibcode : 1981Sci...211..838S . doi : 10.1126/science.211.4484.838 . PMID 17740399 .
- ^ Maureaud, Aurore; Gascuel, Didier; Colléter, Mathieu; Palomares, Maria LD; Du Pontavice, Hubert; Pauly, Daniel; Cheung, William WL (2017). 「海洋食物網の栄養段階機能における地球規模の変化」 . PLOS ONE . 12 (8): e0182826. Bibcode : 2017PLoSO..1282826M . doi : 10.1371/journal.pone.0182826 . PMID 28800358 .
{{cite journal}}: CS1 maint: 記事番号をページ番号として表示 (リンク) - ^ 「海洋酸性化による海洋生物と生態系への影響」報告書OCEANA。2014年12月25日時点のオリジナルよりアーカイブ。 2013年10月13日閲覧。
- ^ 「北極海の酸性化に関する包括的研究」。研究。CICERO。2013年12月10日時点のオリジナルよりアーカイブ。 2013年11月14日閲覧。
- ^ Lischka, S.; Büdenbender J.; Boxhammer T.; Riebesell U. (2011年4月15日). 「海洋酸性化と気温上昇が極地殻翼脚類Limacina helicinaの幼生初期に及ぼす影響:死亡率、殻の劣化、殻の成長」(PDF) .報告書. Biogeosciences. 919– 932ページ . 2013年11月14日閲覧.
- ^ 「南極の海洋生物が脅威にさらされている、と研究で判明」 BBCネイチャー。 2013年10月13日閲覧。
- ^ a b c Tekwa, EW; Watson, James R.; Pinsky, Malin L. (2022). 「温暖化下における種の範囲シフトは、体サイズと食物網相互作用によって媒介される」 . Proceedings of the Royal Society B: Biological Sciences . 289 (1972) 20212755. doi : 10.1098/rspb.2021.2755 . PMC 9006017. PMID 35414233 .
- ^ Chapman, Eric J.; Byron, Carrie J.; Lasley-Rasher, Rachel; Lipsky, Christine; Stevens, Justin R.; Peters, Rebecca (2020). 「気候変動による沿岸生態系の食物網への影響:養殖業への影響」. Marine Environmental Research . 162 105103. Bibcode : 2020MarER.16205103C . doi : 10.1016/j.marenvres.2020.105103 . PMID 33059212 .
- ^「気候変動による海洋と海洋資源への影響」EPA(環境保護庁)
- ^「気候変動指標:海洋生物の分布」EPA(環境保護庁)
- ^ NOAA(アメリカ海洋大気庁)とラトガース大学。2021年。OceanAdapt。2021年1月にアクセス
- ^ Pinsky, Malin L.; Worm, Boris; Fogarty, Michael J.; Sarmiento, Jorge L.; Levin, Simon A. (2013). 「海洋分類群は局所的な気候速度を追跡する」. Science . 341 (6151): 1239– 1242. Bibcode : 2013Sci...341.1239P . doi : 10.1126/science.1239352 . PMID 24031017 .
- ^ Ullah, Hadayet; Nagelkerken, Ivan; Goldenberg, Silvan U.; Fordham, Damien A. (2018年1月9日). Loreau, Michel (編). 「気候変動は栄養段階の変化とシアノバクテリアの増殖を通じて海洋食物網の崩壊を引き起こす可能性がある」 . PLOS Biology . 16 (1) e2003446. doi : 10.1371/journal.pbio.2003446 . ISSN 1545-7885 . PMC 5760012. PMID 29315309 .
- ^ a b気候変動が海洋食物網の崩壊を促すScienceDaily 2018年1月9日
- ^ IUCN (2018) IUCN絶滅危惧種レッドリスト:バージョン2018-1