角竜類

| ケラトサウルス 時間範囲:シネムリアン期-マーストリヒチアン期、 | |||
|---|---|---|---|
 | |||
| 6 種のケラトサウルス類 (左上から右下): ルゴプス、エラフロサウルス、マジュンガサウルス、カルノタウルス、ケラトサウルス、ベルタサウラ | |||
| 科学的分類 | |||
| 王国: | 動物界 | ||
| 門: | 脊索動物 | ||
| クラス: | 爬虫類 | ||
| クレード: | 恐竜 | ||
| クレード: | 竜盤類 | ||
| クレード: | 獣脚類 | ||
| クレード: | 新獣脚類 | ||
| クレード: | アヴェロストラ | ||
| クレード: | †ケラトサウルス・マーシュ、1884 | ||
| タイプ種 | |||
| †ケラトサウルス・ナシコルニス マーシュ、1884年 | |||
| サブグループ | |||
| |||
ケラトサウルスは、鳥類よりもケラトサウルスとの共通祖先がより新しい獣脚類として定義される恐竜のグループであるケラトサウルス亜科に属します。知られている最古のケラトサウルスであるサルトリオヴェナトルは、約1億9900万年前のジュラ紀最初期に遡ります。ケラトサウルス亜科には、ケラトサウルス科、ノアサウルス科、アベリサウルス科の3つの主要なクレードが含まれており、主に(ただし、限定的ではありませんが)南半球で発見されています。もともとケラトサウルス亜科には上記の恐竜に加えて、三畳紀後期からジュラ紀前期のコエロフィソイデア科とディロフォサウルス科が含まれていたため、ケラトサウルスが他の獣脚類からかなり以前に分岐したことが示唆されています。しかし、最近の研究では、コエロフィソイド類とディロフォサウルス類は他のケラトサウルス類と自然なグループを形成しておらず、このグループから除外されていることが示されています。[ 1 ]
角竜類は、1884年にOCマーシュによって記載されたタイプ種、ケラトサウルス・ナシコルニスにちなんで名づけられた。後期ジュラ紀の中型捕食動物、ケラトサウルス・ナシコルニスは最初に発見された角竜類である。ケラトサウルス類は一般に中型だが、より大きなカルノタウルスやかなり小型のノアサウルスなど一部の例外がある。ケラトサウルス類の主な決定的特徴は、装飾が増えた頑丈な頭骨と高さのある頭骨、短い腕である。[ 2 ]これらの特徴は両方とも、アベリサウルス 類などのグループの後期のメンバーで一般的に強調されているが、 C. ナシコルニスなどのより基盤的な種は他の基盤的な獣脚類により似ているように見える。角竜の化石記録は非常に断片化しているため、角竜類の特徴、関係性、初期の歴史は謎に包まれたままであり、激しい議論が続いています。
研究の歴史
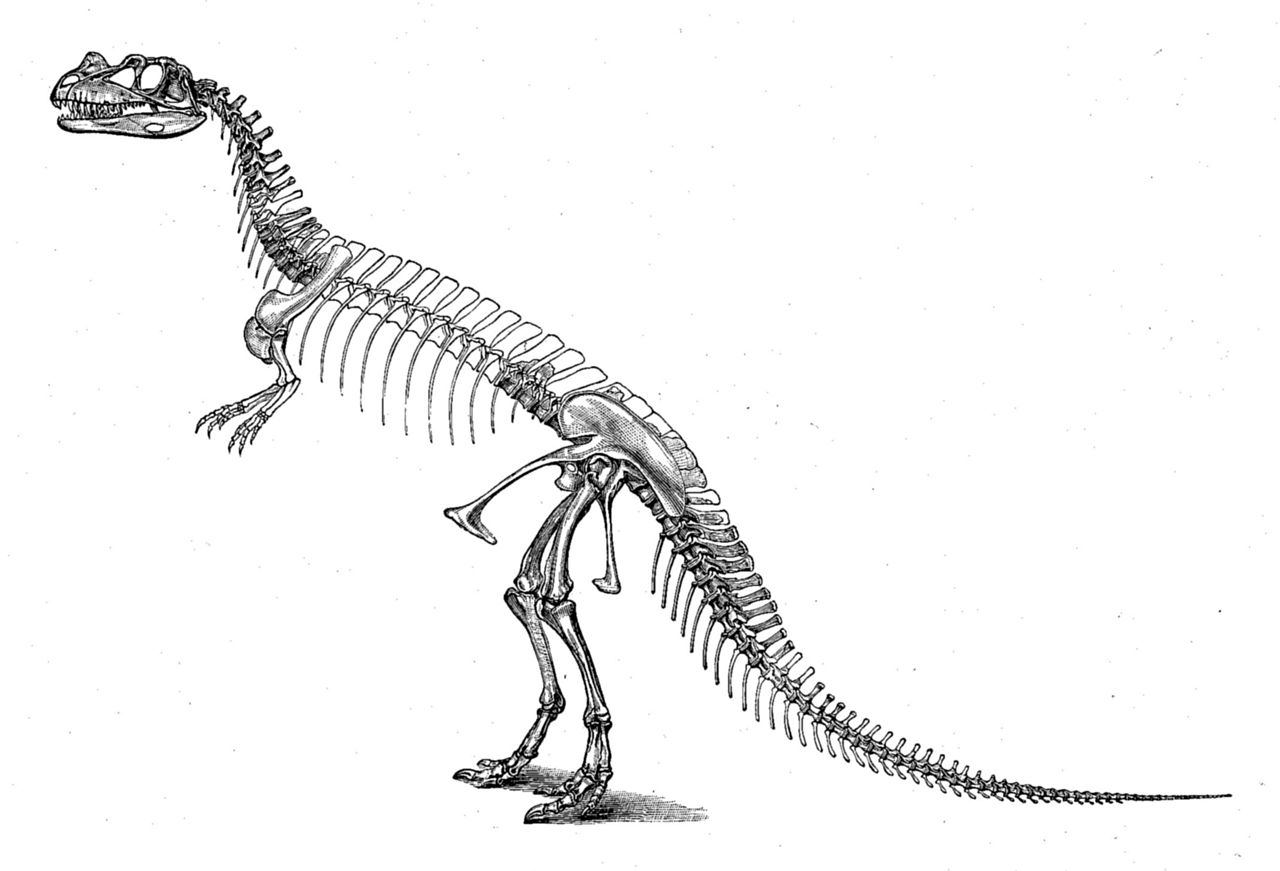

角竜類は1884年、OCマーシュがアメリカ科学誌で初めて記載した。新発見の角竜C. nasicornisについて書いた記事の中で、マーシュはC. nasicornisのしっかりと癒合した中足骨と始祖鳥のそれとの類似点を指摘した。当時発見された恐竜の中でこの特徴を持つのはC. nasicornisだけだったため、マーシュは角竜類は始祖鳥とその関連グループのすぐ近くに位置づけられるべきだと結論付けた。[ 3 ]マーシュはまた、1884年にC. nasicornis を含む 科をCeratosauridaeと命名した。それ以来、主にケラトサウルス属から、他の多くの種がこの科に分類されるようになった。
ケラトサウルス類の説は、マーシュのライバルであるエドワード・ドリンカー・コープによって異論を唱えられた。コープはこの分類群は無効だと主張した。[ 4 ]ケラトサウルス類の説は、30年以上後の1920年にギルモアが賛成を唱えたことで、いくらかの支持を回復した。ギルモアの支持にも関わらず、第一次世界大戦後にこのグループに追加された種はほとんどなく、この説に重点が置かれることもほとんどなかった。実際、20世紀の大半を通じて科学界とケラトサウルス類との最も一般的な関わりは、その存在についての論争であり、ローマー、ラパレント、ラボカット、コルバート、チャリグといった人々によって行われた。
1986年、ジャック・ゴーティエが鳥類の進化を明らかにする試みの中で、獣脚類の大部分を角竜類またはテタヌラ類に分類したことで、角竜類の運命は一変した。彼は角竜類の中に、ケラトサウルス類とコエロフィソイド類を位置づけた。[ 5 ] ゴーティエの論文によって角竜類という用法が再び流行し、1990年代初頭にはアベリサウルス科も角竜類に含まれるようになった。角竜類、コエロフィソイド類、アベリサウルス類の三位一体モデルは、2000年代初頭まで異論なく受け入れられた。2000年代初頭以降、多くの古生物学者がコエロフィソイド類を角竜類から除外し始めた。この見解は、コエロフィソイド類には見られない、ケラトサウルス類とテタヌラ類の間のいくつかの類似点のおかげで、現在では広く受け入れられています。
系統発生

古生物学者の多くは、ケラトサウルス類が三畳紀後期またはジュラ紀初期に他の獣脚類から分岐したと仮説を立てている。しかし、ジュラ紀前期以前にケラトサウルス類は発見されておらず、ジュラ紀中期においても種はまばらである。カラーノやサンプソンをはじめとする多くの科学者は、標本の少なさは当時のケラトサウルス類の豊富さを非難するものではなく、化石記録の乏しさに起因すると仮説を立てている。同様に大きな標本の空白は、特にアベリサウルス科において、下部白亜紀に存在する。近年の発見により、エラフロサウルスと派生した白亜紀ノアサウルス類との関係に関する系統学的結果が様々に分かれている。ケラトサウルスとアベリサウルス類の正確な関係も明確に解明されていない。[ 6 ] [ 7 ] [ 2 ]
現在、ほとんどの古生物学者は、ケラトサウルス類には、ケラトサウルス科とアベリサウルス上科を含む、やや限定的な系統群である新ケラトサウルス類が含まれることに同意しているが、どの分類群が基底的多分岐に位置付けられるかについては多少の差異がある。[ 2 ] [ 8 ]アベリサウルス上科はさらにアベリサウルス科とノアサウルス科に分けられ、アベリサウルス科にはカルノタウリナエ科が含まれる。最近、ラウハットとカラーノはエラフロサウルス科をノアサウルス科に置き、同時に以前のノアサウルス類をノアサウルス科に移した。[ 8 ]彼らは新しいノアサウルス科に、デルタドロメウスとリムサウルスを独自に含めた。
ケラトサウルス科の可能な共形質とケラトサウルスの固有形質を区別することは困難である。なぜなら、関連のあるジェニオデクテスの化石は非常に断片的であるからである。例えば、ケラトサウルスは、吻部の非常に目立つ角によって他のケラトサウルス類と異なる。しかし、ジェニオデクテスは完全な頭骨を伴って発見されておらず、角があったかどうかは不明であるため、角がこのグループに共通の派生的特徴であるとは断定できない。しかし、2 つの属の歯の間には共通の類似点があるため、歯に共形質が認められている。存在する共形質には、口蓋側から見た場合の第 2 および第 3 前上顎骨歯槽の重なり、亜成体/成体の最大歯冠が 6 センチメートルを超えること、側歯の歯冠の 3 分の 2 に四角形以下の近心小歯状部があることなどがある。[ 9 ]現在、ケラトサウルス 属以外で一般的に認知されているケラトサウルス科の種は、白亜紀またはパタゴニアに生息していたジェニオデクテスのみです。エオアベリサウルスとオスタフリカサウルスもケラトサウルス類である可能性は高いですが、ケラトサウルス科に属するかどうかは不明です。[ 10 ] Delcourt (2018) は、ケラトサウルス科を「ケラトサウルス・ナシコルニスを含み、カルノタウルス・サストライを含まない最も包括的な系統群」と定義しました。[ 11 ]

アベリサウルス上科は、多様な角竜類の上位科であり、ケラトサウルス科の姉妹群です。一般的には白亜紀のグループとされていますが、最古のアベリサウルス科の化石はアルゼンチンの中期ジュラ紀( Eoabelisaurus mefiとして分類)と、おそらくマダガスカル(未命名種の断片的な化石)から発見されています。タンザニアの後期ジュラ紀のテンダグル層でも、アベリサウルス科の化石(孤立した左脛骨、右大腿骨、右脛骨)が発見されています。
アベリサウルス上科は白亜紀に南半球で繁栄したが、その起源は少なくとも中期ジュラ紀まで遡ることができ、当時はより広範囲に分布していた(最古のアベリサウルス上科の化石は、約1億7000万年前のオーストラリアと南アメリカの堆積層から発見されている)。 [ 12 ]白亜紀までに、アベリサウルス上科はアジアと北アメリカで絶滅したようで、これはおそらくティラノサウルス上科との競争によるものと思われる。しかし、アベリサウルス科の進化したアベリサウルス上科は、6600万年前の白亜紀-古第三紀絶滅イベントまで南半球に生息し続けた。[ 13 ]
エオアベリサウルスの系統学的位置付けを評価したところ、分析により、この恐竜はアベリサウルス科の中で最も基底的な種であることが判明した。アベリサウルス科の類縁形質には、側方に覆われた涙骨前眼窩、幅広い頸椎前窩、前後方向に短い前尾骨神経棘、前尾骨に腹側溝がないこと、前中尾骨に原始的な中心骨棘板があること、手指骨の遠位歯肉筋が縮小していること、および足の爪骨に屈筋陥凹があることなどがある。エオアベリサウルスの代替系統学的配置は、やや基底的であることを除いて、著しく最適とは言えない。[ 14 ]ノアサウルス科の腕は近縁のアベリサウルス科の腕よりも長く、アベリサウルス科の腕は小さく縮小していた。ノアサウルス科の腕は、進化した鳥類のような獣脚類の腕ほど大きくも特殊化もしていないが、それでも移動や使用が可能で、ノアサウルスなどの大きな爪を持つ属では狩りにも使用できた可能性がある。リムサウルスなどの一部の属は腕と手がやや小型だったが、アベリサウルス科ほど小型ではなかった。ノアサウルス科はまた、機敏で軽く体格が良く、足は長い中央の足骨など、走るのに適応した特徴を示している。ノアサウルス科の大きさは様々で、全長5フィート(1.5メートル)未満の小型のヴェロキサウルスから、全長20フィート(6.1メートル)を超えるエラフロサウルスやデルタドロメウスなどの大型の属までいた。 [ 15 ]

現在知られている最古の角竜はサルトリオヴェナトル・ザネライで、1億9900万~ 1億9700万年前の前期シネムリアンに遡る。[ 16 ]角竜類の起源は北パンゲアにあった可能性があり、そこではサルトリオヴェナトルやその近縁種のベルベロサウルス、カルメロポドゥスの足跡が発見されている。
以下の家系図は、2010年代に行われた様々な研究に基づいて主要な獣脚類グループの関係を統合したもので、獣脚類の中でのケラトサウルス類の位置を示しています。[ 17 ]



.PNG)

.png)
_(Antwerpener_Breiftaube).jpg/1280px-Meyers_grosses_Konversations-Lexikon_-_ein_Nachschlagewerk_des_allgemeinen_Wissens_(1908)_(Antwerpener_Breiftaube).jpg)

以下の系統樹は、 2012年にディエゴ・ポルとオリバー・WM・ラウハットが行った分析に基づくケラトサウルス類の内部関係を示しています。[ 2 ]
| 角竜類 | |
.jpg/1280px-Limusaurus_runner_(flipped).jpg)
.jpg/1280px-Elaphrosaurus_(flipped).jpg)

.jpg/1280px-Masiakasaurus_BW_(flipped).jpg)


.png)


.jpg/1280px-Ilokelesia_(flipped).jpg)


.png)

2017年に発表されたリムサウルスの個体発生に関する論文では、異なる結論が導かれた。他の解析とは異なり、ノアサウリダエ科はケラトサウルスよりも基底的な位置に置かれ、ケラトサウルスは定義上アベリサウルス科に属するとされた。[ 18 ]この結論は後に、2018年に発表されたケラトサウルスの古生物学に関する論文でさらに詳しく述べられ、アベリサウルス科とケラトサウルス科を含む新しい系統群、エトリガンスアウルアが命名された。[ 7 ]以下の系統樹は、論文で示された最新の系統樹のコンセンサス樹である。
| ケラトサウルス上科 | |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_(Antwerpener_Breiftaube).jpg){kind=link}
.jpg){kind=link}
.jpg){kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
古生物学
解剖学
_2_(15415407082).jpg){kind=link}
_2_(15415407082).jpg/1280px-Masiakasaurus_knopfleri_theropod_dinosaur_(Maevarana_Formation,_Upper_Cretaceous;_near_Berivotra,_northern_Madagascar)_2_(15415407082).jpg)
角竜類の特徴として、頭骨の増大と装飾、前肢の短縮が挙げられます。同様に、角竜類は腸骨、坐骨、恥骨、および距骨と踵骨が癒合しています。[ 4 ] C. nasicornis など、このグループ内で派生度の低い種では、頭骨の隆起や前肢の短縮といった特徴はそれほど顕著ではありませんでした。 C. nasicornisの頭骨は基底獣脚類の型にかなり似ており、特徴的な鼻の隆起と、同時代のアロサウルスに似た涙骨の隆起がありました。C. nasicornis の歯はアロサウルスよりも大きく、一部の古生物学者は、より大きな獲物を攻撃するのは困難だったと仮定しています。しかし、アベリサウルス類はこれらの特徴の多くを極限まで押し進めていた。ほとんどのアベリサウルス類は前肢が大きく短縮しており、カルノタウルスはどの大型獣脚類よりも前肢を縮小させていた。[ 19 ]ポール・セレノは、新たに発見されたルゴプス・プリムス の特徴を分析した後、これらのアベリサウルス類の特徴の多くが腐肉食に適していた可能性があると仮説を立てた。[ 20 ] 体長が大幅に縮小したにもかかわらず、ケラトサウルス類のどの分類群も指や前肢の重要な要素を失ったことはなかった。ケラトサウルス類には関節の変異も見られ、他の大型獣脚類よりも肩の可動性が優れていたのではないかと仮説が立てられている。[ 1 ]
ケラトサウルスの歯には2つの種類が知られています。1つは縦溝を持つ歯、もう1つはエナメル質に脈状の溝を持つ歯です。どちらの種類の歯も、冠は涙滴形の断面を持ち、中央に隆起部が走っています。歯の基部の断面は口内の位置によって異なり、前歯はより対称性が低い断面をしています。
完全な骨格が記載されているのは、最も進化したアベリサウルス科(カルノタウルスやアウカサウルスなど)のみであるため、科全体の骨格の特徴を確定するのはより困難である。しかし、ほとんどの骨格は少なくとも一部の頭骨から知られているため、既知の共通の特徴は主に頭骨から得られる。アベリサウルス科の頭骨の特徴の多くはカルカロドントサウルス科と共有されている。[ 21 ]これらの共通の特徴と、アベリサウルス科が南アメリカでカルカロドントサウルス科に取って代わったと思われる事実から、2つのグループは関連があるとの説が浮上した。ノアサウルス科は、足の第2指に独特の「鎌状爪」を持つ、特徴的なアベリサウルス上科であると考えられており、これはデイノニコサウルス科のものと収斂的に発達した。ノアサウルス科のうち、アルゼンチンのノアサウルス属(白亜紀後期)とリガブエイノ属(白亜紀前期)は、不完全な標本から知られており、その中には分離した非爪骨や、ノアサウルスには鉤爪が含まれていた。これらの要素を詳細に調査した結果、第2ペダル指の捕食鉤爪と推定されるものは実際には手指の第1指または第2指に属し、また、推定される非爪骨、あるいは両属も手指に属することが示唆された。[ 22 ]
古生態学
地理
ケラトサウルスはジュラ紀前期までに世界中に広く分布していたと考えられています。しかし、白亜紀にはローラシア大陸からほぼ姿を消したと考えられており、発見された少数の標本はゴンドワナ大陸から再導入された可能性があります。[ 23 ]白亜紀の北アメリカでは、ケラトサウルスの標本は確認されていません。
特にアベリサウルス科はゴンドワナ大陸、とりわけ白亜紀に大きな繁栄を遂げました。ゴンドワナ大陸とローラシア大陸の標本が最近発見され、後期ジュラ紀、あるいは中期ジュラ紀のものと年代測定されました。これにより、アベリサウルス科の年代は大幅に延長されました。一部の古生物学者は、広大な砂漠がアベリサウルス科を後期ジュラ紀まで南ゴンワナ大陸に閉じ込めていた可能性があると仮説を立てています。[ 2 ]相関関係か因果関係かはさておき、後期白亜紀のケラトサウルス類は、基盤的テタヌラ類(アフリカ)やコエルロサウルス類(北米およびアジア)が優勢な地域ではあまり発見されていないことが広く観察されています。
ダイエット
{kind=link}

ほとんどの獣脚類と同様に、ケラトサウルス類は肉食だった。ただし、リムサウルスやベルタサウラなどの一部のノアサウルスは、歯のない嘴を持つ雑食性または草食性だった。[ 24 ] [ 25 ]ケラトサウルスは大量の魚や他の水生生物を食べていたと主張されてきたが、多くの古生物学者によって異論が出ている。[ 26 ]アロサウルス などの大型動物の歯型は、ケラトサウルスが腐肉食を頻繁に行っていたことを示している。 [ 27 ]アベリサウルス科 の興味深い顎は、さまざまな食事の予測を引き起こしている。カルノタウルスに関するある研究では、その短い頭骨のおかげで、その噛みつきは素早いが比較的弱い噛みつきにより、小さな獲物を狩るのに適していたことがわかった。[ 28 ] 一方、他の古生物学者のグループは、カルノタウルスの噛みつきが比較的強力で、大型の獲物を狩って傷つけるのに適していたことを発見しました。[ 29 ]
頭骨は腐肉食のために作られたという説もある。ケラトサウルスの食性に関する議論は、特に近年のアベリサウルスの発見増加に伴い、非常に活発である。歯列ベースのデータマトリックスを用いた系統分類学的分析、および獣脚類の歯の測定値の大規模データセットを用いた判別分析とクラスター分析という3つの手法を用いることで、アベリサウルス科獣脚類に確実に分類できる3つの歯の形態型を特定した。これらの形態型が異なるアベリサウルス科の亜系統群を表しているのか、それとも同じアベリサウルス科の顎の中で異なる位置の実体を表しているのかは不明である。しかしながら、このような特定は、アベリサウルス科が竜脚類の死骸を食べていたというさらなる証拠を提供するものである。[ 30 ]
マジュンガサウルスに関する研究によると、この恐竜は他の獣脚類よりも成長が遅く、成体になるまでに約20年かかったことが示されている。他のアベリサウルス科の属に関する同様の研究では、このゆっくりとした成長はアベリサウルス科全体に共通する特徴であった可能性があることが示されている。ノアサウルス亜科は、南アメリカ、マダガスカル、インドなどの南の大陸と島でのみ知られている後期白亜紀のノアサウルス亜科である。エラフロサウルス亜科は、小さな頭骨と長い首と脚を持つ、軽量な獣脚類であった。リムサウルスが何らかの指標であるならば、成体のエラフロサウルス亜科にはまったく歯がなく、口はおそらく角質の嘴で縁取られていた。リムサウルスの成長した標本に草食恐竜のものと似た胃石や化学的特徴が保存されていることから、リムサウルスなどのエラフロサウルス亜科は成体になると主に草食であった可能性が高い。
古環境

モリソン層とテンダグル層で発見されたことから、ケラトサウルス科は他の大型捕食動物と共存していたことが分かります。北米では、C. nasicornisなどのケラトサウルス科の種は、当時この地域に広く生息していた竜脚類などの食物を巡って、アロサウルス科(A. fragilis )と競合していたと考えられます。アフリカとヨーロッパでも、ケラトサウルス科の種は同様の食物源を巡って他の大型捕食動物と競合していました。クリーブランド・ロイド恐竜採石場でC. nasicornisが複数のアロサウルス科の化石とともに発見されたことは、ケラトサウルス科の種と他の捕食動物がどれほど密接に共存していたかを示す良い例です。
アベリサウルス上科の角竜のほとんどはマダガスカル、アジア、時にはアフリカで発見されています。アベリサウルス科は白亜紀に古代の南方超大陸ゴンドワナで繁栄し、今日ではその化石は現代のアフリカ大陸と南アメリカ大陸、インド亜大陸、マダガスカル島で発見されています。マダガスカルでは、フランスの古生物学者シャルル・デペレがマジュンガサウルスを発見しました。マジュンガサウルスは私たちが知る最も一般的なアベリサウルス上科です。南アメリカでは、スコルピオヴェナトル、キルメサウルス、アウカサルス、イロケレシア、ピコネモサウルスなど多くのアベリサウルス上科が知られています。クルピ・イタアタは、マリリア層(バウル層群、バウル盆地)で初めて正式に命名された脊椎動物であり、バウル盆地のマーストリヒチアンにおける数少ない獣脚類の記録の一つである。骨盤と前尾椎の解剖学的構造に基づき、アベリサウルス科との類縁関係は十分に確立されているが、他のアベリサウルス科とのより密接な関係は依然として不明である。この標本は、ブラジルの化石記録ではまだよく知られていないアベリサウルス上科と、南米におけるこのグループの獣脚類恐竜の分布について新たな情報を提供する。[ 31 ]これらの発見は、アベリサウルス上科が生息していた地域で最も一般的な大型捕食恐竜であったことを示している。[ 32 ]
参照
参考文献
- ^ a b Carrano, Matthew T.; Sampson, Scott D. (2008-01-01). 「角竜類(恐竜類:獣脚類)の系統発生」. Journal of Systematic Palaeontology . 6 (2): 183– 236. Bibcode : 2008JSPal...6..183C . doi : 10.1017/S1477201907002246 . ISSN 1477-2019 . S2CID 30068953 .
- ^ a b c d e Diego Pol & Oliver WM Rauhut (2012). 「パタゴニア産中期ジュラ紀アベリサウルス類と獣脚類恐竜の初期の多様化」 . Proceedings of the Royal Society B: Biological Sciences . 279 (1804): 3170–5 . doi : 10.1098/rspb.2012.0660 . PMC 3385738. PMID 22628475 .
- ^ Marsh, OC (1884). 「ケラトサウルスの結合した中足骨について」 . American Journal of Science . s3-28 (164): 161– 162. Bibcode : 1884AmJS...28..161M . doi : 10.2475/ajs.s3-28.164.161 . S2CID 131737380. 2020年6月24日時点のオリジナルよりアーカイブ。2019年12月3日閲覧。
- ^ a bワイシャンペル, デイビッド・B.; ピーター, ドッドソン; オスモルスカ, ハルシュカ (2007). 『恐竜』 . カリフォルニア大学出版局. ISBN 978-0-520-24209-8. OCLC 493366196 .
- ^ 「竜盤類の単系統性と鳥類の起源」カリフォルニア科学アカデミー紀要8巻1986年。ISSN 0885-4629。 2014年11月3日時点のオリジナルよりアーカイブ。2017年5月31日閲覧。
- ^カウ、A. (2024)。「捕食恐竜の大進化のための統一フレームワーク」。ボッレッティーノ デッラ ソシエタ 古生物イタリアーナ。63 (1): 1-19 .
- ^ a b Delcourt, Rafael (2018-06-27). 「ケラトサウルスの古生物学:南方支配者の進化と生態に関する新たな知見」 . Scientific Reports . 8 (1) 9730. Bibcode : 2018NatSR...8.9730D . doi : 10.1038/s41598-018-28154 - x . ISSN 2045-2322 . PMC 6021374. PMID 29950661 .
- ^ a bラウハット、オリバー WM;カラーノ、マシュー T. (2016-11-01)。 「タンザニア、テンダグルのジュラ紀後期の獣脚類恐竜エラフロサウルス・バンベルギ」。リンネ協会の動物学雑誌。178 (3): 546–610 .土井: 10.1111/zoj.12425。ISSN 0024-4082。
- ^ Hendrickx, Christophe; Mateus, Octávio; Araújo, Ricardo; Choiniere, Jonah (2019-11-24). 「非鳥類獣脚類恐竜の歯の特徴の分布:分類学的可能性、同形性の程度、そして主要な進化的傾向」 . Palaeontologia Electronica . 22 (3): 1– 110. doi : 10.26879/820 . hdl : 10362/98436 . ISSN 1094-8074 . 2021年11月8日時点のオリジナルよりアーカイブ。 2024年6月17日閲覧。
- ^ Rauhut, Oliver WM (2011). 「テンダグル(タンザニア)のジュラ紀後期の獣脚類恐竜」.古生物学特別論文集. 86 : 195–239 .
- ^ Delcourt (2018). 「ケラトサウルスの古生物学:南方支配者の進化と生態に関する新たな知見」 . Scientific Reports . 8 (1) 9730. Bibcode : 2018NatSR...8.9730D . doi : 10.1038/s41598-018-28154-x . PMC 6021374. PMID 29950661 .
- ^ David B. Weishampel; Peter Dodson; Halszka Osmólska (2004年11月6日). 『恐竜:第2版』 . カリフォルニア大学出版局. p. 109. ISBN 978-0-520-24209-8。
- ^ Martín D. Ezcurra, MD and Agnolín, FL (2012). 「ローラシア中期ジュラ紀のアベリサウルス上科恐竜と獣脚類の古生物地理学および進化への影響」 Proceedings of the Geologists' Association(オンライン先行公開)
- ^ T カラーノ、マシュー。 「マダガスカルの白亜紀後期のマジュンガサウルス クレナティッシムス(獣脚類: アベリサウルス科)の付属肢骨格」 。リサーチゲート。
- ^ Pol, D.; Rauhut, OW (2012). 「パタゴニア産中期ジュラ紀アベリサウルス類と獣脚類恐竜の初期の多様化」 . Proceedings . Biological Sciences . 279 (1741): 3170– 3175. doi : 10.1098/rspb.2012.0660 . PMC 3385738. PMID 22628475 .
- ^ Sasso, Cristiano Dal; Maganuco, Simone; Cau, Andrea (2018-12-19). 「イタリアのジュラ紀前期に発見された最古の角竜類(恐竜類:獣脚類)が、鳥類の3本指の手の進化に光を当てる」 . PeerJ . 6 e5976 . doi : 10.7717/peerj.5976 . ISSN 2167-8359 . PMC 6304160. PMID 30588396 .
- ^ Hendrickx, C.; Hartman, SA; Mateus, O. (2015). 「非鳥類獣脚類の発見と分類の概要」PalArch's Journal of Vertebrate Palaeontology . 12 (1): 1– 73.
- ^ Wang, S.; Stiegler, J.; Amiot, R.; Wang, X.; Du, G.-H.; Clark, JM; Xu, X. (2017). 「ケラトサウルス類獣脚類における極端な個体発生的変化」(PDF) . Current Biology . 27 (1): 144– 148. Bibcode : 2017CBio...27..144W . doi : 10.1016/j.cub.2016.10.043 . PMID 28017609 .
- ^ボナパルト, ホセ; ノバス, フェルナンド; コリア, ロドルフォ (1990). 「Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia. Contributions in Science」(PDF) .ロサンゼルス郡立自然史博物館. 416 : 1– 42. 2016年6月29日時点のオリジナル(PDF)からアーカイブ。 2017年5月31日閲覧。
- ^ Sereno, Paul C.; Brusatte, Stephen L. (2008). 「ニジェール、下部白亜紀エルラズ層産の基底アベリサウルス科およびカルカロドントサウルス科獣脚類」 . Acta Palaeontologica Polonica . 53 (1): 15– 46. doi : 10.4202/app.2008.0102 . hdl : 20.500.11820/5d55ed2e-52f2-4e4a-9ca1-fd1732f2f964 .
- ^ T カラーノ、マシュー。「マダガスカルの白亜紀後期のマジュンガサウルス クレナティッシムス (獣脚類: アベリサウルス科) の付属肢骨格」。 2007 年 6 月。
- ^アグノリン、フェデリコ;キアレッリ、パブロ。「ノアサウルダ科(恐竜:アベリサウルス上科)の爪の位置とアベリサウルス類の手骨の進化への影響」ResearchGate。
- ^トルトーサ、ティエリー;エリック・バフェフォー。ヴィアル、ニコラ。デュトゥール、イヴ。エリック・トゥリニ。ジル・シェラン (2014)。 「白亜紀後期の南フランスから発見された新しいアベリサウルス類の恐竜:古生物地理学的意義」。古生物学の記録。100 (1): 63–86。ビブコード: 2014AnPal.100...63T。土井:10.1016/j.annpal.2013.10.003。
- ^ Xu, Xing; Clark, James M.; Mo, Jinyou; Choiniere, Jonah; Forster, Catherine A.; Erickson, Gregory M.; Hone, David WE; Sullivan, Corwin; Eberth, David A. (2009-06-18). 「中国産ジュラ紀ケラトサウルスが鳥類の指の相同性を明らかにする」 ( PDF) . Nature . 459 (7249): 940– 944. Bibcode : 2009Natur.459..940X . doi : 10.1038/nature08124 . ISSN 0028-0836 . PMID 19536256. S2CID 4358448. 2017年9月22日時点のオリジナルよりアーカイブ( PDF) . 2019年9月6日閲覧。
- ^ de Souza GA、Soares MB、Weinschütz LC、Wilner E、Lopes RT、de Araújo OM、Kellner AW (2021)。「南米初の無歯ケラトサウルス類」。科学的報告書。11 (1) 22281。Bibcode : 2021NatSR..1122281D。土井: 10.1038/s41598-021-01312-4。PMC 8602317。PMID 34795306。
- ^ J., Currie, Philip; B., Koppelhus, Eva; A., Shugar, Martin; L., Wright, Joanna (2004). 『羽毛のあるドラゴン:恐竜から鳥類への移行に関する研究』インディアナ大学出版局. ISBN 978-0-253-34373-4. OCLC 895411495 .
{{cite book}}: CS1 maint: 複数の名前: 著者リスト (リンク) - ^ 「北米ジュラ紀後期における捕食性恐竜による獲物の骨の利用、および中生代における恐竜による獲物の骨の利用に関するコメント(PDFダウンロード可能)」ResearchGate。2017年5月31日閲覧。
- ^マゼッタ、ヘラルド V.シシリーノ、アドリアン P.ブランコ、R. アーネスト。カルボ、ネストル (2009-09-12)。 「角のある肉食恐竜カルノタウルス・サストライの頭蓋力学と機能的解釈」。脊椎動物古生物学のジャーナル。29 (3): 822–830。ビブコード: 2009JVPal..29..822M。土井: 10.1671/039.029.0313。hdl : 11336/34937。ISSN 0272-4634。S2CID 84565615。
- ^ケネス・カーペンター (2005). 『肉食恐竜』 インディアナ大学出版局. ISBN 978-0-253-34539-4. OCLC 895411496 .
- ^ G. Meso, Jorge; Hendrickx, Christophe; A. Baiano, Mattia; I. Canale, Juan; Salgado, Leonardo; Diaz Martinez, Ignacio. 「アルゼンチン、パタゴニア、リオネグロ州後期白亜紀アレン層の竜脚類骨格に付随する孤立した獣脚類の歯」Acta Paleontologica Polonica . 2021年6月13日時点のオリジナルよりアーカイブ。2021年6月30日閲覧。
- ^ A. Farke, Andrew (2013). "「マダガスカルのチューロニアン期のアベリサウルス上科獣脚類恐竜」" . PLOS ONE . 8 (4) e62047 . Bibcode : 2013PLoSO...862047F . doi : 10.1371/journal.pone.0062047 . PMC 3630149 . PMID 23637961 .
- ^メンデス、アリエル; E・ノバス、フェルナンド;ヴィドイ・イオリ、ファビアーノ。「ブラジル、サンパウロ州、バウル盆地(白亜紀後期)からのアベリサウルス科獣脚類の新記録」。リサーチゲート。
外部リンク
