ディオスコレア・アラタ
| ディオスコレア・アラタ | |
|---|---|
 | |
| 紫芋の断面 | |
| 科学的分類 | |
| 王国: | 植物界 |
| クレード: | 維管束植物 |
| クレード: | 被子植物 |
| クレード: | 単子葉植物 |
| 注文: | ヤマノイモ目 |
| 家族: | ヤマノイモ科 |
| 属: | ヤマノイモ |
| 種: | D. アラタ |
| 二名法名 | |
| ディオスコレア・アラタ | |
| 同義語[ 2 ] | |
リスト
| |
ディオスコレア・アラタ(Dioscorea alata)は、ウベ( / ˈ uː b ɛ , - b eɪ /)、ウビ、パープルヤム、オオヤムなど、様々な別名で 呼ばれるヤムイモ(塊茎)の一種である。塊茎は通常、鮮やかな紫から明るいラベンダー色(これが一般名の由来)だが、クリーム白から無地の白のものがある。サトイモや沖縄のサツマイモである紅芋( Ipomoea batatas 'アヤムラサキ' )と混同されることもあるが、ディオスコレア・アラタは沖縄でも栽培されている。原産地はアジアとオセアニアの熱帯地方である。 [ 3 ]品種によっては大型になるものもある。1939年頃、パプアニューギニアの東セピック地方マプリクで栽培された「マンバタップ」と呼ばれるオオヤムイモは、長さが3.5メートル(11フィート)もあった。 [ 4 ] [ 5 ]
名前
D. alataはアジア、特にフィリピンを原産地とし、南米の熱帯地域、そしてアメリカ合衆国南東部を経て帰化しているため、これらの地域では様々な名前で呼ばれています。英語だけでも、パープルヤムのほか、テン・マンス・ヤム(ten-months yam)、ウォーターヤム(water yam)、ホワイトヤム(white yam)、ウィングドヤム(winged yam)、バイオレットヤム(violet yam)、ガイアナ・アロールート(Guyana arrowroot)、あるいは単にヤム(yam)などの一般的な名前があります。[ 3 ]
 | この記事には、野生近縁種(D. nummularia、D. transverse、D. hastifolia;祖先不明)に関する情報が不足しています[doi:10.1111/boj.12374]。(2021年10月) |
栽培の歴史

ディオスコレア・アラタ(Dioscorea alata)は、オーストロネシア文化において最も重要な主食作物の一つである。東南アジア島嶼部とニューギニアでは、デンプン質の塊茎を目的に、独自に栽培・栽培されていた様々なヤムイモの一種であり、丸いヤムイモ(Dioscorea bulbifera )、ウビガドゥン(Dioscorea hispida)、小ヤムイモ(Dioscorea esculenta)、太平洋ヤムイモ(Dioscorea nummularia)、五葉ヤムイモ(Dioscorea pentaphylla)、ペンシルヤム(Dioscorea transversa)などが含まれる。[ 6 ]これらのうち、D. alataとD. esculentaだけが定期的に栽培・食用とされていたが、残りは毒素ジオスコリンの含有量が高く、正しく調理する必要があるため、飢餓時の食料とみなされていた。 [ 7 ] D. alataはD. esculentaよりも多く栽培されていますが、これは主に塊茎が大きいためです。[ 8 ]
D. alataとD. esculentaは、オーストロネシア船による長距離輸送に最も適しており、オーストロネシア人の進出範囲のほぼ全域に渡って運ばれました。特にD. alataは太平洋諸島とニュージーランドに導入されました。また、オーストロネシア人の航海者によってマダガスカル島とコモロ諸島にも持ち込まれました。[ 9 ] [ 10 ] [ 11 ]

ムラサキイモの原産地はフィリピンですが、考古学的証拠によると、オーストロネシア人の進出以前には東南アジア島嶼部とニューギニアでも利用されていたことが示唆されています。ムラサキイモは真の栽培種であると考えられており、栽培種のみが知られています。栽培品種の大部分は不妊であるため、島嶼部への導入は純粋に人間の行為によるものと限定されており、人類の移動を示す良い指標となっています。一部の研究者は証拠なしに東南アジア大陸部起源説を唱えていますが、フィリピンとニューギニアでは表現型の多様性が最も顕著です。[ 12 ] [ 13 ] [ 14 ]
クック湿地遺跡における初期の農地跡と植物遺骸の考古学的証拠に基づき、著者らは、この植物がニューギニア高地で約1万年前から栽培化され、約4000年前頃にラピタ文化を経て、 D. nummulariaやD. bulbiferaと共に東南アジア島嶼部に広まったと示唆している。一方、D. esculentaはラピタ文化によってニューギニアにもたらされたと考えられている。この時期には、オーストロネシア人との接触による革新、特に湿地耕作の発展によって農業革命がもたらされたという証拠もある。[ 15 ] [ 16 ]
しかし、ボルネオ島のニア洞窟 (後期更新世、4万年前未満)とパラワン島のイレ洞窟(約1万1千年前)からは、おそらくD. alataと思われるはるかに古い遺骨が、食用に加工する必要がある有毒植物ウビガドゥン(D. hispida )の遺骨とともに発見されている。これは栽培を証明するものではないが、人類がすでにデンプン質植物を利用する知識を持っていたこと、そしてD. alataが東南アジア島嶼部原産であったことを示している。さらに、D. alataが真の種なのか、それとも一般に考えられているよりもはるかに古くから栽培されていたのかという疑問も浮上する。[ 6 ] [ 17 ] [ 18 ] [ 19 ] [ 20 ] [ 21 ]
紫ヤムは東南アジア、特にフィリピンにおいて重要な作物であり、鮮やかな紫色の品種は様々な伝統的および現代的なデザートに広く利用されています。メラネシアでも重要な作物であり、収穫時の塊茎の大きさと結びついた儀式用としても栽培されています。しかし、東ポリネシアとニュージーランドでは、サツマイモなどの他の作物が導入されて以来、その重要性は薄れつつあります。[ 8 ]
用途
料理
紫ヤムイモには食用の塊茎があり、ほんのり甘く、土っぽく、ナッツのような味がして、サツマイモやタロイモを思わせる。特に紫色の栽培品種は、アントシアニンの含有量が多いため、料理を独特の鮮やかな紫色に変えることができる。[ 22 ]紫ヤムイモは、加工できるデンプンも高く評価されている。 [ 3 ] 紫ヤムイモはフィリピン料理で最も一般的であり(ウベまたはウビと呼ばれている)、ウベケーキ、ウベチーズケーキ、ウベクリンクルなど多くのフィリピンのデザートに広く使われているほか、アイスクリーム、ミルク、ドーナツ、タルト、ジャム、その他のペストリーの材料や風味としても使われている。茹でたり、焼いたり、ウベハラヤと呼ばれる甘いデザートとして食べられることが多い。[ 23 ] [ 24 ] [ 25 ]紫イモを使ったデザートは、近年フィリピン料理を通じて「ウベ」というフィリピン語名でアメリカに伝わってきました。デザートに鮮やかな紫色を与えることから、特に人気があります。[ 22 ] [ 23 ] [ 26 ]
紫イモは、色、味、料理への応用が似ていることから、紫色/すみれ色のサツマイモとよく混同されます。しかし、他のヤムイモと同様に、紫イモはサツマイモよりもしっとりとした食感です。また、紫イモはサツマイモよりもアントシアニン含有量が多いです。その他の点では、ほとんどのレシピで両者は同じように使用できます。[ 27 ] [ 28 ]
- フィリピンのウベハラヤ(紫芋のピューレ)
- フィリピンのウベ・チャンポラード
- フィリピン産のウベとムスコバドのピアヤ
- ココナッツウベソースをかけた沖縄風サツマイモパンケーキ
- ウベクレープケーキ一切れ
- フランをトッピングした紫芋のカップケーキ
- フィリピンのウベドーナツ
- ウベ・マモン


_01.jpg/1280px-Piaya_flatbread_(Philippines)_01.jpg)






サプリメント、民間療法とその副作用
栄養補助食品として入手可能で民間療法にも使用されているが、 D. alataに治療効果があるという臨床的証拠はない。[ 29 ] D. alataサプリメントの使用は、エストロゲン、抗凝固薬を服用している人や妊娠中および授乳中の人には悪影響を及ぼす可能性がある。 [ 29 ] D. alataサプリメントの使用にアレルギー反応を起こす人もいる。[ 29 ]
D. alataは比較的高いレベルのシュウ酸(486~781 mg/100 g乾燥物)を含み、[ 30 ]これは抗栄養作用と腎臓結石の形成に関連しています。[ 31 ]
その他の用途
紫色の品種の色は、様々なアントシアニン色素によるものです。[ 32 ]この色素は水溶性であり、食品着色料としての可能性があると提案されています。[ 33 ] D. alataは、観賞価値のために庭で栽培されることもあります。[ 3 ]
侵略的外来種として
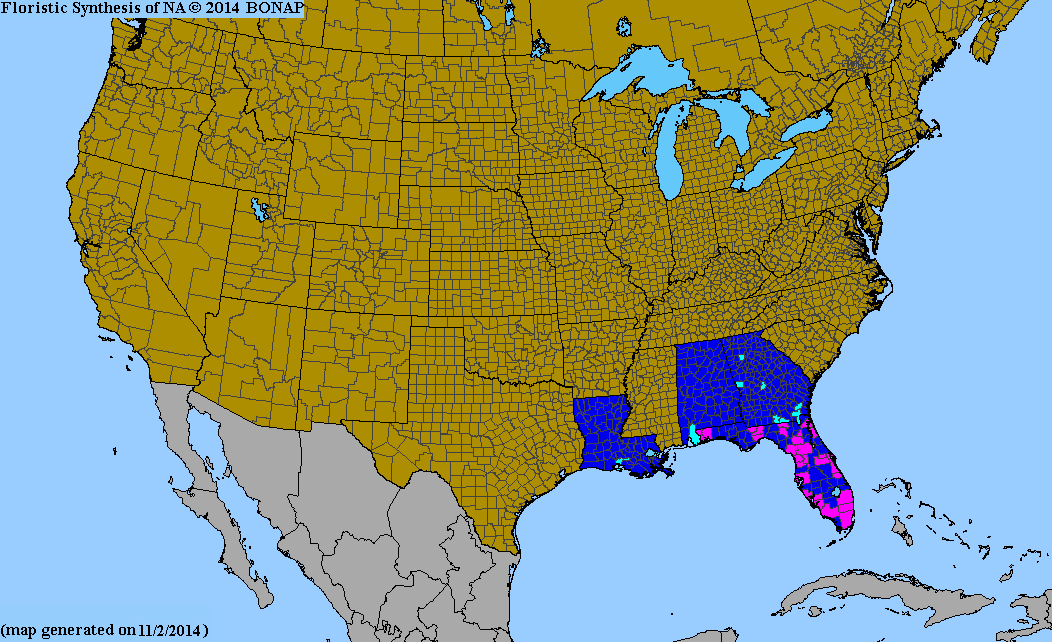
ディオスコレア・アラタはフィリピンとその周辺地域(台湾と日本の琉球諸島)が原産です。原産地から多くの野生化し、中国南部および中東部、アフリカ、マダガスカル、西半球、インド洋および太平洋の様々な島々に帰化しています。[ 34 ] [ 35 ]野生下ではハイチ、アメリカ合衆国、ハワイ、ルイジアナ、ジョージア、アラバマ、プエルトリコ、米領バージン諸島、そしてフロリダでも生息しており、フロリダでは侵略的外来種とみなされています。[ 36 ] [ 37 ]
ギャラリー
- 紫芋の葉
- 紫芋の花

.jpg/1280px-Starr-061106-1437-Dioscorea_alata-flowers-Maui_Nui_Botanical_Garden-Maui_(24572888200).jpg)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_01.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
参照
参考文献
- ^ Dioscorea alata は、 Species Plantarum 2: 1033で初めて記載・出版されました。 1753年。 「Name – Dioscorea alata L.」Tropicos .ミズーリ州セントルイス:ミズーリ植物園. 2011年5月26日閲覧。
- ^ “The Plant List: A Working List of All Plant Species” . 2024年5月13日時点のオリジナルよりアーカイブ。 2023年11月4日閲覧。
- ^ a b c d「Dioscorea alata」。遺伝資源情報ネットワーク。米国農務省農業研究局。 2011年5月26日閲覧。
- ^ Lea D (1966年3月). 「マプリク地域でのヤムイモ栽培」.パプアニューギニア農業ジャーナル. 18 (1): 4 脚注.
- ^ピーターソンC(1986年10月)「国家の誕生」ナショナルジオグラフィック170 ( 4):482。
- ^ a b Barker G, Hunt C, Barton H, et al. (2017年8月). 「ボルネオの『文化的な熱帯雨林』」(PDF) . Quaternary International . 448 : 44– 61. Bibcode : 2017QuInt.448...44B . doi : 10.1016/j.quaint.2016.08.018 .
- ^ Bevacqua RF (1994). 「東南アジアにおける園芸の起源とポリネシア航海者による太平洋諸島への栽培植物の拡散:ハワイ諸島の事例研究」(PDF) . HortScience . 29 (11): 1226– 1229. doi : 10.21273/HORTSCI.29.11.1226 .
- ^ a b “*クフィ~うひ、うひ” .テ マラ レオ: ランゲージ ガーデン。ベントン・ファミリー・トラスト。2019 年1 月 21 日に取得。
- ^ Crowther A, Lucas L, Helm R, 他 (2016年6月14日). 「古代の作物はオーストロネシア語族の西方への拡大を示す最初の考古学的証拠となる」 . Proceedings of the National Academy of Sciences . 113 (24): 6635– 6640. Bibcode : 2016PNAS..113.6635C . doi : 10.1073/pnas.1522714113 . PMC 4914162. PMID 27247383 .
- ^ Beaujard P (2011年8月). 「マダガスカルへの最初の移住者と植物の導入:言語学的・民族学的証拠」(PDF) .アザニア:アフリカの考古学研究. 46 (2): 169– 189. doi : 10.1080/0067270X.2011.580142 . S2CID 55763047 .
- ^ウォルター A、レボット V (2007).オセアニアの庭園。 IRD エディション - CIRAD。ISBN 978-1-86320-470-5。
- ^マラパ R、アルナウ G、ノワイエ J、他。 (2005 年 11 月)。 「AFLP マーカーで明らかにされた、ヤムイモ (Dioscorea alata L.) の遺伝的多様性と D. numularia Lam. および D. transversa Br. との関連性」。遺伝資源と作物の進化。52 (7): 919–929。ビブコード: 2005GRCEv..52..919M。土井: 10.1007/s10722-003-6122-5。S2CID 35381460。
- ^ Cruz V, Altoveros N, Mendioro M, et al. (1999). 「フィリピンの食用ヤムイモコレクションにおける多様性の地理的パターン」. Plant Genetic Resources Newsletter . 119 : 7–11 .
- ^ Paz VJ (1999). 「新石器時代の東南アジア島嶼部への人類の移動:考古植物学的証拠の探求」インド太平洋先史協会紀要. 18 (Melaka Papers Vol. 2): 151– 158. doi : 10.7152/bippa.v18i0.11710 (2025年7月12日現在非アクティブ).
{{cite journal}}: CS1 maint: DOI inactive as of July 2025 (link) - ^ Chaïr H, Traore RE, Duval MF, et al. (2016年6月17日). 「サトイモ( Colocasia esculenta (L.) Schott)の遺伝的多様化と分散」 . PLOS ONE . 11 (6) e0157712. Bibcode : 2016PLoSO..1157712C . doi : 10.1371/ journal.pone.0157712 . PMC 4912093. PMID 27314588 .
- ^ Bayliss-Smith T, Golson J, Hughes P (2017). 「フェーズ4:主要な処分場、溝のような溝、そして格子状の畑」 . Golson J, Denham T, Hughes P, Swadling P, Muke J (編).パプアニューギニア高地クック湿地における1万年の耕作. terra australis. 第46巻. ANU Press. pp. 239– 268. ISBN 978-1-76046-116-4。
- ^ Barker G, Lloyd-Smith L, Barton H, et al. (2011). 「ボルネオ島サラワク州ニア洞窟における採餌から農耕への移行」 . Antiquity . 85 (328): 492– 509. doi : 10.1017/S0003598X00067909 . S2CID 131187696 .
- ^ Balbaligo Y (2007年11月15日). 「フィリピン、パラワン島、イレ洞窟における2007年の発掘調査に関する簡潔な報告」 .考古学研究所論文集. 18 (2007): 161. doi : 10.5334/pia.308 .
- ^ Barton H (2005). 「熱帯雨林の食料採集民の事例:サラワク州ニア洞窟におけるデンプン記録」(PDF) . Asian Perspectives . 44 (1): 56– 72. doi : 10.1353/asi.2005.0005 . hdl : 10125/17222 . S2CID 11596542 .
- ^ Barton H, Denham T (2011). 「島嶼東南アジアとメラネシアにおける先史時代の菜食と社会生活」(PDF) . Barker G, Janowski M (編). 「なぜ耕作するのか?東南アジアにおける採食・農耕の変遷への人類学的・考古学的アプローチ」 . マクドナルド考古学研究所. pp. 61– 74. ISBN 978-1-902937-58-8. 2019年3月28日時点のオリジナル(PDF)からアーカイブ。2019年1月21日閲覧。
- ^ Reynolds T, Barker G, Barton H, et al. (2013). 「ニアにおける最初の現代人、約5万~3万5千年前」(PDF) . Barker G (編).東南アジア島嶼部における熱帯雨林での採食と農業. マクドナルド考古学研究所. pp. 133– 170. ISBN 978-1-902937-54-0。
- ^ a b Sutherlin M. 「紫芋ウベについて知っておくべきことすべて」Chowhound . 2019年5月18日閲覧。
- ^ a b Kearns L. 「ウベは食べ物を完璧な紫色に変える天然成分」ハフィントン・ポスト。 2019年5月18日閲覧。
- ^ Bueno A. 「当たり前だと思っていたウベ、料理の宝石よ、万歳」 CNNフィリピン2019年11月1日時点のオリジナルよりアーカイブ。 2019年5月18日閲覧。
- ^ Valdeavilla R (2018年6月6日). 「Ube: The Philippine Purple Yam (More Popular Than Vanilla!)」 . culture trip . 2019年5月18日閲覧。
- ^ 「ガーニーのドーナツショップがユニークな作りたてドーナツを販売」 ABC7シカゴ、2016年11月11日。
- ^ 「Ubeかそうでないか、それが問題だ…そしてFrieda'sが答える」 Frieda 's 2017年4月17日. 2019年5月18日閲覧。
- ^ 「注目の食材:デザートの材料となる紫芋、ウベ」 OneGreenPlanet 、 2017年3月24日。 2019年5月18日閲覧。
- ^ a b c「ワイルドヤム」 . Drugs.com. 2020年7月24日. 2021年3月15日閲覧。
- ^ Wanasundera JP, Ravindran G (1994). 「ヤムイモ( Dioscorea alata )塊茎の栄養評価」. Plant Foods Hum Nutr . 46 (1): 33– 9. Bibcode : 1994PFHN...46...33W . doi : 10.1007/ BF01088459 . PMID 7971785. S2CID 36080604 .
- ^ Otegbayo BO, Oguniyan DJ, Olunlade BA, et al. (2017年11月1日). 「ナイジェリア産ヤムイモ(Dioscorea spp.)在来種の生化学組成、抗栄養性、ミネラルのバイオアベイラビリティにおける遺伝子型変異の特徴づけ」 . Journal of Food Science and Technology . 55 (1): 205– 216. doi : 10.1007 / s13197-017-2913-0 . ISSN 0022-1155 . PMC 5756202. PMID 29358812 .
- ^ Moriya C, Hosoya T, Agawa S, et al. (2015). 「紫イモ由来の新規アシル化アントシアニンとその抗酸化活性」 . Biosci Biotechnol Biochem . 79 (9): 1484–92 . doi : 10.1080/ 09168451.2015.1027652 . PMID 25848974. S2CID 11221328 .
- ^ Li J, Zhang L, Liu Y (2013). 「応答曲面法による紫芋からの天然色素抽出の最適化とその安定性」 . Journal of Chemistry . 2013 : 1–5 . doi : 10.1155/2013/590512 .
- ^キュー植物園指定植物科チェックリスト
- ^中国植物誌、第24巻、296ページ、shen shu、Dioscorea alata Linnaeus、Sp. Pl. 2: 1033。1753。
- ^ 「 Dioscorea alata(ウォーターヤム)のプロファイル」PLANTSデータベース。USDA 、NRCS 。2011年5月26日閲覧。
- ^北米生物相プログラム、2013年郡分布図
{kind=link}
外部リンク
- 「Dioscorea alata」 .統合分類情報システム. 2006年2月18日閲覧。
- 2012年3月20日、Wayback Machineにアーカイブされた画像。Forestry Images(ジョージア大学侵入種・生態系健全性センターのウェブページ)より
